Taxonomía
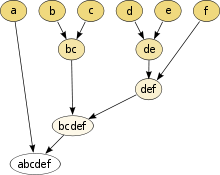
Árbol filogenético, en este caso en forma de dendrograma[nota 1] (cada grupo, incluidos los de los nodos, es un grupo de organismos), como cada nodo del dendrograma se corresponde con un clado, entonces es un cladograma. Cada clado agrupa a todos los descendientes de una población ancestral común, es decir es un grupo monofilético. Los nodos terminales (aquí simbolizados por letras individuales) no pueden ir más allá de las especies, ya que por definición, por debajo de la categoría especie las poblaciones no evolucionan como linajes independientes.

Árbol filogenético (en este caso, las líneas representan la secuencia de organismos ancestros y descendientes, es un filograma[cita 1]) que muestra un ejemplo de diversificación de una especie ancestral en tres especies presentes en la actualidad. Cada nodo del árbol (como el marcado con un círculo rojo) representa la divergencia de un linaje ancestral en dos linajes que evolucionan independientemente, esto es, que tienen el potencial de adquirir su propio set de cambios evolutivos. En barras rojas se muestran las apomorfías: los nuevos estados del carácter. Un cladograma se puede interpretar como una secuencia de ancestros y descendientes, lo cual es una hipótesis que se superpone al dendrograma obtenido al determinar las apomorfías que agrupan los linajes vivientes. No todos los investigadores están de acuerdo en interpretar un cladograma de esta forma, porque por ejemplo la historia filogenética del grupo puede no haberse dado en forma de cladogénesis.[cita 2]

Del árbol filogenético se puede derivar una clasificación de los organismos. Los taxones terminales se circunscriben y agrupan en taxones de rango sucesivamente más alto. Los anidamientos deben tener congruencia o "consistencia lógica" con el árbol filogenético, aunque la forma en que se tiene que dar esta consistencia está en discusión.[14] Nótese que la clasificación tiene forma de dendrograma.
La Taxonomía (del griego τάξις táxis ‘ordenamiento’ y νόμος nómos ‘norma’ o ‘regla’) es, en su sentido más general, la ciencia de la clasificación. Habitualmente se emplea el término para designar a la Taxonomía Biológica, la «teoría y práctica de clasificar organismos».[15]
[cita 3][cita 5] Como se la entiende en la actualidad, la clasificación biológica tiene que ser congruente con las hipótesis de árbol filogenético disponibles, en ella, los organismos se agrupan en taxones mutuamente excluyentes a su vez agrupados en taxones de rango más alto también mutuamente excluyentes, de forma que cada organismo pertenece a uno y sólo un taxón en cada rango o "categoría taxonómica". En cada taxón, los organismos poseen caracteres (singular carácter) cuyos estados del carácter los diferencian de los organismos fuera del taxón, y son los atributos que en conjunto se utilizan para delimitar los taxones biológicos. La mayoría[cita 6] de los especialistas ve a las especies, a las que ubican en taxones en la categoría taxonómica del mismo nombre, como una realidad objetiva, y a los diferentes conceptos de especie como diferentes aproximaciones para reconocer los linajes que evolucionan independientemente, por lo que a pesar de las discrepancias entre autores en su concepto taxonómico y por lo tanto en general en los límites taxonómicos de cada una, suele ser considerada la categoría taxonómica más importante[cita 7][cita 8] en la clasificación. Los taxones supraespecíficos son los que agrupan especies y se jerarquizan en categorías taxonómicas de rango cada vez más alto: género (grupo de especies), familia, etc. hasta el reino. En esta área también está debatido el concepto de taxón a utilizar. Muchos especialistas afirman que sólo la "filosofía taxonómica" para agrupar los organismos en taxones de la escuela cladista logra que la clasificación ofrezca el servicio que se espera de ella al resto de las ramas de la biología, pero la escuela evolucionista tiene razones para delimitar con otros conceptos los taxones, también congruentes con las hipótesis de árbol filogenético, y las dos escuelas se atribuyen la creación de sistemas de clasificación que cumplen en mayor medida sus funciones de sistemas de almacenamiento y recuperación de datos, y de predictores del estado de los caracteres y las direcciones de la evolución que no se han medido.
La Taxonomía también se ocupa de debatir y actualizar los Códigos Internacionales de Nomenclatura, de los que por razones históricas hay uno para cada disciplina (Zoología, Botánica y Bacteriología), cuya utilización para reglamentar el uso de nombres "formales" está consensuada desde hace unos 100 años. Un Código define el sistema taxonómico —los Códigos en uso definen el recién descripto, conocido como el "sistema lineano", del que se deriva la definición de taxón más abajo descripta—; decide cómo se forman los nombres y cuál es el "tipo" con el que se heredan de taxón en taxón, y explicita sus Principios de Nomenclatura, cuya validez está por encima de las reglas, con el objetivo o "principio básico"[19] de proveer la máxima estabilidad en la nomenclatura. Los Códigos en uso no evitan las "diferencias filosóficas" entre taxónomos en los conceptos taxonómicos a utilizar ni en la categoría especie ni en las categorías supraespecíficas que son una fuente de inestabilidad frecuente en los nombres de los taxones.
La Taxonomía Biológica es aquí tratada como una subdisciplina de la Biología Sistemática, que además tiene como objetivo la reconstrucción de la filogenia, o historia evolutiva, de la vida.[cita 4] Como aquí comprendida la Taxonomía abarca la decisión de qué sistema taxonómico utilizar, la de qué conceptos taxonómicos asociar a esos taxones, la delimitación de esos taxones dejando explícitos los métodos que fueron utilizados para alcanzarla, la formalización de sus nombres en la subdisciplina de la Nomenclatura, y también las herramientas para la Determinación o identificación de especímenes.[cita 4][cita 7] Como se calcula que sólo un 10% de las especies está descripto, esas descripciones tienen diferentes grados de resolución,[cita 9] y las especies evolucionan (cambian con el tiempo en términos de sus caracteres) y cambia su distribución, la determinación funciona recursivamente con la descripción y catalogación de especies en el área llamada alfa-Taxonomía o Taxonomía Descriptiva, o, deslindándola de la definición minimalista de Taxonomía que se maneja desde hace décadas y debido a que se intergradan (se van indiferenciando a medida que el foco de atención se traslada de una a la otra) en esa categoría taxonómica, el área de la Sistemática Descriptiva[23] o Ciencia de la Biodiversidad,[cita 7] la que se ocupa del descubrimiento, la identificación, la descripción, la clasificación y la catalogación[cita 7] de la biodiversidad del planeta, utilizando el organismo como unidad de trabajo y cuya prioridad[24] es la descripción de todas las especies dentro de un contexto evolutivo.[cita 7]
La Historia de la Taxonomía ha sido largamente recorrida por éstos y otros interrogantes y a lo largo de ella fueron formándose el sistema de clasificación y las reglas que nos han quedado como legado.
La crisis de biodiversidad que es foco de atención desde mediados de 1990, la cantidad de datos acumulados con los análisis de ADN, y la posibilidad de intercambiar información a través de internet, han revitalizado a esta ciencia en el ambiente científico desde las décadas de 1990-2000, y han generado un debate acerca de la necesidad de hacer reformas sustanciales a los Códigos, algunas de las cuales aún se están debatiendo y otras ya se han incorporado. Algunas de estas propuestas son el BioCode, el PhyloCode, el agregado de atributos al tipo nomenclatural como podrían ser los nuevos "marcadores de ADN" (ADN barcodes), y las relacionadas con la informatización de los datos y la utilización formal de Internet.
Índice
1 Definición de taxón
2 La taxonomía según las diferentes escuelas
3 Las dos funciones de los sistemas de clasificación
4 Construyendo una clasificación
5 La clasificación en la categoría especie
6 Nomenclatura biológica
6.1 Categoría taxonómica
6.2 Nombre científico
6.3 ¿Por qué cambian los nombres de los taxones?
7 Determinación o identificación de especímenes
7.1 La colecta
7.2 La identificación (Determinación)
8 Historia de la taxonomía
8.1 De la taxonomía linneana al origen de los Códigos
8.2 Taxonomía y evolución
8.3 Formalización de las normas: los Códigos
8.4 Historia reciente
8.5 Relación con las taxonomías populares
9 Perspectivas actuales de la ciencia de la Taxonomía
9.1 El impedimento taxonómico
9.2 El rol del ADN y los "marcadores de ADN"
9.3 La crisis de biodiversidad
9.4 Informática de la Biodiversidad
9.5 La armonización de los Códigos y el BioCode
9.6 Propuestas de un registro central de nombres
9.7 El PhyloCode
10 Véase también
11 Notas
12 Citas
13 Referencias citadas
14 Bibliografía
15 Enlaces externos
Definición de taxón
El taxón es un grupo de organismos con circunscripción, posición y rango. De la estructura de la clasificación biológica como dada en la introducción, comúnmente llamada "taxonomía linneana" o "sistema linneano" (pero ver[cita 10]), se deriva la definición de taxón, que como explicitan los Códigos de Nomenclatura,[cita 12] es un grupo de organismos que posee:
circunscripción: el conjunto de caracteres obtenidos de los organismos que lo componen que, junto con la regla de decisión propuesta al observalos, diagnostican al taxón, establecen sus límites o lo delimitan, vuelven al taxón único, diferencian sus organismos claramente de los organismos fuera del taxón; la circunscipción se define en función de poder determinar si un organismo nuevo pertenece o no al taxón mediante la observación de sus caracteres según el protocolo dado, y ella puede incluir idealmente o no una lista de caracteres diagnósticos (presentes en ese taxón y sólo en ese taxón), el término aristotélico[cita 11] "definición" se sigue utilizando en el Código de Zoología para referirse a la descripción exhaustiva que debería hacerse de un taxón al nombrarlo, de la que se extraería la circunscripción;[cita 13]- un rango o jerarquía (absoluta)[nota 2] llamado categoría taxonómica;
- y una posición taxonómica formada por los taxones de rango más alto que lo contienen.
El taxón se redefine según el "sistema" taxonómico del que se deriva, si bien la circunscripción es necesaria para ubicar organismos nuevos en su respectivo taxón y por lo tanto, fundamental en todas las clasificaciones. Así, puede ser modificada su definición para adaptarse a taxonomías que no utilizan rangos, o pueden definirse los taxones de formas más restringidas, como en las clasificaciones que usan otros atributos además de los caracteres para circunscribir sus taxones, como la localidad, pero las filosofías taxonómicas que han formado tres escuelas de la sistemática que dejaron un legado a la materia, se diferencian en el concepto que debe utilizarse para aglomerar los organismos en taxones, es decir su definición conceptual, su delimitación conceptual, el concepto que relaciona los organismos del taxón más entre sí que con los demás organismos, los diferentes "conceptos de taxón" o "conceptos taxonómicos".[cita 14]
La taxonomía según las diferentes escuelas

Un clado[cita 1] en una definición más rigurosa es el conjunto de organismos en las poblaciones actuales (en naranja) que se hipotetiza que son todas las descendientes de una secuencia de ancestros y descendientes con ancestro común (en naranja y blanco). Pero se puede ser más riguroso todavía[cita 2].
Según la escuela cladista, la que predomina hoy en día,[cita 16] la taxonomía es la ciencia que debe decidir qué clados del árbol filogenético serán representados por taxones y en qué categoría taxonómica ubicarlos.[cita 17]
Si bien en los libros de texto el clado en general[18] se define como "una secuencia de una población ancestral y todos sus descendientes", resulta ser una definición poco rigurosa:[cita 1] el clado, como lo utilizan los taxónomos, agrupa sólo poblaciones vivientes o al menos las que se utilizaron para tomar los datos, las que en el método original de Hennig —el fundador,[40] o la "voz más articulada"[38] del cladismo— se asocian como descendientes de un mismo linaje cuando se "encuentran" o determinan sus apomorfías: los cambios de carácter en el linaje ancestral.[cita 1] Otras formas de análisis filogenético como los métodos fenéticos (cuyo punto de partida es ciego en lo que respecta a la evolución de los caracteres[cita 18][cita 19]) fueron incorporados a la definición de clado, de forma que pueden hipotetizar un "clado" (poblaciones agrupadas por poseer caracteres compartidos desde el ancestro común más reciente) mediante métodos de análisis no Hennigianos.[cita 1] El lector interesado puede recurrir a las siguientes lecturas, listadas en Judd et al. (2007[18]): Farris (1979[53][5]), Humphries y Parenti (1986[54]), Brooks y McLennan (1991[55]), Forey et al. (1992[56]), Cracraft y Donoghue (2004[57]).
"Clado" proviene de cladogénesis.[cita 1] El conjunto de todos los descendientes de un linaje se llama grupo monofilético (mono: único, phylum: linaje) sensu Hennig,[cita 15] y los términos usualmente son utilizados como sinónimos (pero ver[cita 2]). Los taxones que representan grupos monofiléticos se llaman "taxones monofiléticos", y las apomorfías del clado que representan, que pueden haberse encontrado, no alcanzan normalmente como circunscripción. Las apomorfías pueden haberse perdido o vuelto a derivar en algunos miembros del grupo, pueden presentar variación fenotípica en diferentes ambientes en los que se presenta el grupo que debe ser expresada, pueden presentar los mismos genes pero otras variaciones debido a interacciones con otros genes en diferentes individuos o grupos que también debe ser expresada, y pueden ser compartidas por otros clados que las hubieran adquirido independientemente. Sólo después de haber obtenido una circunscripción (con apomorfías con toda su variación fenotípica y genotípica, y las plesiomorfías necesarias) y decidido una categoría taxonómica, se puede considerar al grupo un taxón "lineano" y decidir si nombrarlo según las reglas formales de nomenclatura.
 La Taxonomía en la actualidad. La Taxonomía (definida según la escuela cladista) decide qué clados[cita 1] serán los conceptos taxonómicos representados por taxones con su circunscripción, posición taxonómica, categoría taxonómica, y nombre formal o científico. |
Hay otras escuelas de clasificación. Quizás la más importante, dentro de las "minoritarias"[cita 16] es la que considera que los grupos parafiléticos también deberían ser representados por taxones nombrados formalmente, si los grupos que los conforman son lo suficientemente similares entre sí y lo suficientemente disímiles del clado que queda afuera (escuela evolucionista[cita 20]). Los autores listados en Judd et al. 2007[18] para el lector interesado: Simpson 1961,[15] Ashlock 1979,[61][63] Ejemplos de "taxones parafiléticos" que se utilizan en las "clasificaciones evolucionistas" son los procariotas (parafiléticos con respecto a los eucariotas) en el clásico sistema de clasificación en reinos iniciado por Whittaker (1969[64] – Margulis 1971[65]) y que hoy se sigue utilizando (p.ej. Cavalier-Smith 1998[cita 21]), y las dicotiledóneas, parafiléticas con respecto a las monocotiledóneas (p.ej. Takhtajan 2009[66]).
 Comparación de árboles filogenéticos (filogramas) de grupo monofilético, grupo parafilético y grupo polifilético. Dibujado a partir de Cavalier-Smith (2010[40]). |

La simplificación que sufrió el cuerpo de la planta al haberse asociado con hongos para su desarrollo hizo creer durante mucho tiempo que las psilotáceas (foto) habrían retenido muchos caracteres ancestrales similares a los encontrados en las primeras plantas vasculares del registro fósil como Rhynia, aunque no se habían encontrado fósiles que sostuvieran este argumento; y hasta mediados de los '2000 se las había ubicado como grupo hermano de ese grupo. Las reconstrucciones filogenéticas para las que el enorme número de caracteres tomados de las líneas de evidencia moleculares (de ADN) fueron decisivos consensuaron que su ubicación filogenética estaba anidada entre los helechos y afines como grupo hermano de las ofioglosáceas, lo que obligó a reinterpretar el origen evolutivo del grupo.
Otra escuela que fue encabezada por RR Sokal y PHA Sneath[67] (Sokal y Sneath 1963,[47] Sneath y Sokal 1973[68][2][67]) recordaba que como no había forma de testear si eran verdaderas las "hipótesis previas" sobre evolución, los razonamientos se hacían circulares: los "grupos preferidos" determinan las "apomorfías preferidas" que a su vez determinan los "grupos preferidos".[cita 22] En los '60 cada autor de una clasificación proponía un árbol completamente distinto[cita 23] y eran pocos los grupos que se mantenían entre autores. Sokal y Sneath no solo presentaban novedosos métodos de análisis "numéricos"[nota 3] –cuantitativos– que sacaban provecho de las en ese momento novedosas computadoras, sino que también eran métodos "fenéticos"[nota 3]: intentaban no asumir hipótesis de evolución para armar el árbol, que debía buscarse siempre que existiera la posibilidad.[cita 18][cita 24] En las últimas décadas, en que "toda la sistemática es en alguna forma numérica"[67] debido a la cantidad de caracteres que se obtienen a su vez de cantidad de líneas de evidencia diferentes (a las tradicionales se sumaron los análisis moleculares de ADN, los caracteres bioquímicos y de ultraestructura), se pudo dar vida útil a los métodos de esta escuela fenética que al mismo tiempo perdió visibilidad, porque a medida que se agregan hipótesis evolutivas para aumentar la congruencia entre líneas de evidencia diferentes, los taxónomos van llegando a un consenso acerca de cuáles serán las "verdaderas" de ellas con las que estimar la filogenia, con las que inferir las relaciones cladísticas.[cita 24] Desde mediados de los '90 la sistemática vivió momentos emocionantes: se empezaba a vislumbrar un único árbol predictivo y robusto, y efectivamente, muchos grupos fueron abandonados por no estar basados en hipótesis evolutivas correctas, incluidos algunos de larga data que causaron mucha sorpresa. Los autores de esta tradicionalmente llamada "escuela"[cita 20] enfatizaban que sólo propusieron métodos de análisis numérico,[4] que fueron solo una parte de importantes avances numéricos en análisis filogenético, sistemática molecular, análisis morfométrico e identificación numérica.[67]
Hoy, las clasificaciones cladistas predominan por sobre las evolucionistas.[cita 16] Las dos escuelas admiten que se necesitan nombres tanto para los grupos parafiléticos como para los monofiléticos que los abarcan, la discusión entre cladistas y evolucionistas radica en cuál es el sistema de nombres que debería formalizarse en una clasificación.[cita 25] Los evolucionistas formalizan nombres de grupos parafiléticos, y mencionan con nombres informales los grupos monofiléticos no clasificados (por ejemplo Cavalier-Smith 1998 en adelante[cita 21] formaliza los reinos y nombra los supergrupos de eucariotas sin formalizarlos), mientras que los cladistas formalizan sólo grupos monofiléticos, llamando con nombres informales los grupos parafiléticos (por ejemplo Adl et al. 2005[87] formalizan los supergrupos de eucariotas y proponen el nombre informal procariotas o bacterias para ese grupo parafilético).
Las dos funciones de los sistemas de clasificación
Más allá de los resultados de los análisis filogenéticos, todas las escuelas coinciden en que la formalización de taxones en sistemas de clasificación tiene dos funciones, en miras de ser un servicio para todo un abanico de usuarios finales, que no siempre pueden hacerse coincidir:
- Deben permitir hacer identificaciones,[18][cita 26] o visto de otra manera, deben funcionar como un sistema de almacenamiento y recuperación de datos; para lo que cada taxón debe estar claramente delimitado: un espécimen encontrado del que se pudieran obtener los estados de sus caracteres, debería poder ubicarse inequívocamente en un taxón conocido. Los científicos en todas las disciplinas deben utilizar los taxones como unidad de trabajo, y publicar los resultados de sus trabajos en relación al taxón estudiado. Por lo tanto los nombres científicos de los organismos, los taxones formales, deben representar el consenso de la comunidad científica, sólo de esta forma funcionan como llave de acceso a un cuerpo de información que refiere siempre al mismo grupo de organismos, aunque esté disperso en libros y revistas escritos en muchas lenguas por diferentes autores, o proceda de diferentes campos de la biología o la divulgación: como idealmente hacen referencia los taxónomos, "El nombre de un organismo es la llave hacia su literatura".[cita 27]
- Deben permitir hacer predicciones[18][cita 26], o pronósticos (que a diferencia de las predicciones no están ligados a un método sino a un autor, en este caso el autor de la clasificación y quizás también el autor final) y dar explicaciones. Por ejemplo, cuando se encuentra un compuesto de interés médico en una planta, es común que haya interés en investigar si ese compuesto se encuentra también en otras especies relacionadas con ella; una taxonomía bien armada debería permitirle al investigador presuponer a priori los límites de su investigación para los que sea más probable confirmar sus hipótesis y ahorrar esfuerzos de muestreo.[18]
La decisión de si armar la clasificación para predecir o para identificar recorrió toda la historia de la taxonomía; las dos funciones coincidirían en una clasificación que fuera al mismo tiempo "natural" como "artificial",[cita 28] es decir cuyos conceptos taxonómicos sean "naturales" y cuyos taxones sean "artificiales" —como siempre lo son, en las definiciones aquí manejadas—, con claves de identificación que la reensamblen en grupos con parecidos superficiales y por lo tanto muy poco predictivos, pero con reglas de decisión explícitas con las que se llegue con facilidad al taxón terminal.[cita 26] La dificultad radica en que si se hace el ejercicio de buscar un solo carácter que sea diagnóstico de un taxón (si el organismo posee un estado del carácter, pertenece al taxón, si posee otro estado, no pertenece al taxón, invariablemente, es decir una clasificación monotética), se notará que esta búsqueda puede fallar al menos potencialmente en todos los grupos "naturales", debido a que todos los caracteres son inestables en algún grado y pueden evolucionar, y otros pueden presentarse como polimorfismos y el mismo estado coalescer de forma de encontrarse en parte de taxones diferentes, y esta situación también se encuentra si se toma un conjunto de caracteres. La forma alternativa de circunscripción es proponer los estados de forma probabilística, esto es, un organismo es miembro de un taxón si posee un número muy alto de estados de caracteres similares a la descripción del taxón, especialmente de los poco lábiles.[cita 29] El mismo algoritmo politético dará siempre el mismo resultado, basado en las probabilidades, pero a veces puede no ser el esperado. Aquí se encuentra la diferencia fundamental entre lo que los autores llaman clasificaciones "naturales" y "artificiales": en la clasificación natural, la determinación es una probabilidad de estar acertado, dicho de otra manera un porcentaje de veces se espera que la determinación llegue al taxón correcto pero no al concepto taxonómico esperado. Esto es porque el grupo (el concepto taxonómico) existe independientemente de si el taxónomo lo circunscribió correctamente, e independientemente de la clave de identificación utilizada. En las clasificaciones artificiales, con tanto taxones como conceptos taxonómicos artificiales, la determinación, aunque provenga de un pesaje probabilístico de los caracteres, siempre es la correcta, aunque el grupo no resulte ser lo que el taxónomo tenía en mente. El trabajo del taxónomo se manifiesta en su habilidad para corregir los grupos artificiales creados por el hombre (los taxones) de forma de acercarlos cada vez más a los grupos naturales (los conceptos "naturales") con cada agregado de información nueva.[nota 4]
Si un grupo "natural" es un grupo "predictivo" y esto, en taxones superiores a especie, lo logran el número y peso de los caracteres, medidos y no medidos, compartidos por los miembros del grupo que evolucionan juntos, entonces la "naturalidad" de los grupos será una cuestión de grado, y habrá grupos más o menos similares, o más o menos "naturales".[cita 28] Su componente más importante es el histórico o filogenético,[2] que además ofrece impresas en sus similitudes fenéticas las restricciones construccionales que restringen las direcciones de la evolución del grupo de una forma que no puede ser explicada por factores ecológicos.[40] Tanto cladistas como evolucionistas se adjudican las clasificaciones con mayor valor predictivo, o "más naturales" (p.ej. T Cavalier-Smith[cita 30], E Mayr[cita 31], NI Platnick[cita 32], Hörandl y Stuessy[cita 33]).
Construyendo una clasificación
Hoy en día los taxónomos, aún dentro de la escuela cladista (p. ej. Judd et al. 2007[18]:33), tienen claro que no es de utilidad que cada clado o concepto de taxón por arriba de especie se formalice, es decir se corresponda con un "taxón nombrado" acorde a las reglas consensuadas en los Códigos Internacionales de Nomenclatura. Las decisiones a la hora de sintetizar el árbol, inevitables, pueden parecer un poco arbitrarias por falta de métodos confiables para lograr los dos objetivos recién mencionados[cita 34], pero se pueden generalizar algunos criterios ([18]:32-35):

Un taxón es congruente con todas las líneas de evidencia cuando es posible derivarlo de todas ellas. En el ejemplo diagramado, en el que los árboles son interpretados como filogramas hennigianos (sin marcar las apomorfías en cada rama), todos los taxones parafiléticos posibles derivados de los dos árboles también hubieran podido derivarse del árbol consensuado, ejemplos más complejos en los que esto no es posible entran en juego cuando se tienen en cuenta las apomorfías.
- El criterio principal es la fuerza de la evidencia que respalda al grupo[18]: para convertir un grupo en taxón formal debe haber un análisis filogenético que lo respalde de forma robusta, sobre la base de más de una línea de evidencia de forma que todas ellas muestren congruencia[nota 5]. Según Judd et al.[18] esto se evidencia en una cantidad importante de apomorfías que definan su ancestro.
- Debe haber una cantidad de caracteres diagnósticos que permitan diferenciarlo claramente del resto de los taxones.[18] Según algunos sistemáticos cada taxón debería poder ser diagnosticado morfológicamente en lo posible (p.ej.[18],[101]).
- Deberían poder inferirse aspectos de su biología únicos a ese taxón, para ello debería tener caracteres morfológicos interesantes ("obvios") que permitan que el taxón se comporte como una unidad predictiva (Judd et al. 2007[18]:33).
- Algunos autores[18] afirman que los taxones deberían tener, en lo posible, entre tres y siete subtaxones, un número que puede manejar con facilidad la memoria humana.[102] En las palabras de Davis y Heywood (1963[88][18]): "Debemos ser capaces de ubicar a los taxones en taxones de categoría más alta de forma que podamos encontrarlos de nuevo". Los sistemas de clasificación permiten la creación de categorías intermedias hasta lograr el número de taxones deseados en cada una.
- Por estabilidad de la nomenclatura, los taxones que ya han sido nombrados en el pasado deben continuar con el mismo nombre en lo posible, a menos que haya fuertes justificaciones para abandonarlos.
Hay dos criterios para decidir la categoría taxonómica. Uno, el tradicional, es el que existe desde los inicios de la clasificación y es anterior a la teoría de la evolución; éste decide la categoría taxonómica según el grado de variación del plan corporal entre taxones.[40] Desde la incorporación de la teoría de la evolución al pensamiento científico, se interpreta a la categoría taxonómica como la que deriva de un grupo de apomorfías que aportan cierto grado de variación en el plan corporal del taxón en relación a los demás taxones en la misma categoría.[40] Entonces los reinos y los filos (las categorías más altas) derivan de transformaciones evolutivas importantes que definieron de nuevo el plan corporal, a las que siguió un batallón de novedades evolutivas forzadas a adaptarse al nuevo cambio, mientras que las categorías más bajas, como géneros y especies, poseen variaciones en caracteres más triviales, y por lo tanto más inestables.[40] Esta construcción de categorías, basada en variación de caracteres más a menos estables, es la que llegó hasta nuestros días y probó ser útil para los usuarios finales de una clasificación (ej. Cavalier-Smith 2010[40]). Otro criterio de clasificación en categorías fue el iniciado por Hennig (1950,[36] 1966[37]), quien proponía que la edad geológica absoluta de un grupo fuera el criterio para decidir su categoría taxonómica, y si bien hay científicos trabajando para lograr esta meta en algunos grupos[103] según otros parece ser un concepto abandonado[nota 6]. Las categorías resultaron ser útiles para los objetivos de la clasificación (ej.[105][106],[107]), pero algunos autores las desestiman por considerarlas arbitrarias[nota 7] y hasta los hay que proponen eliminar el sistema de clasificación con categorías (por ejemplo en de Queiroz 2006[nota 8]). Por razones históricas el sistema definido por los Códigos en uso utiliza las categorías linneanas de clasificación: reino, filo o división, clase, orden, familia, género y especie.
La clasificación en la categoría especie

Filograma de tipo "spindle" de los vertebrados. Los linajes se caracterizan por ser "cohesivos" en cada momento y "continuos" en el tiempo, su pool genético está integrado pero cambia a lo largo del tiempo, por lo que no necesariamente representan similitud fenotípica ni genotípica (Simpson 1961,[15] Mayr 1963,[110] Ghiselin 1974,[111] Hull 1976,[112] 1978,[113] Wiley 1978,[114] Hennig 1966,[37] 1975,[115] Platnick 1977[116]:97, como citados en Hull 1979[3]).
Un "concepto de taxón" particular es el de especie. La mayoría de los especialistas ve las especies, a las que ubican en taxones en el rango o categoría taxonómica del mismo nombre, como un "taxón natural", una realidad objetiva,[cita 6] haciendo uso los diferentes conceptos de especie de las diferentes aproximaciones o las diferentes propiedades emergentes en el nivel de organización del linaje que evoluciona independientemente, esto es que, al menos potencialmente, adquiere su propio set de cambios evolutivos. A diferencia de la situación en los rangos más altos, el hecho de que este rango es objetivo y que existe al menos entre o en forma de dos cotas superior e inferior es poco cuestionado en la práctica, le es intuitivo al lego,[117] y es el concepto que hace que cobren sentido objetivos de otras ramas como "conservar todas las especies": un linaje que se pierde no puede ser regenerado sin cambios evolutivos en linajes emparentados. Pero como no se sabe cómo van a evolucionar los linajes y por lo tanto cuáles de las delimitaciones posibles constituyen una "unidad evolutiva" en proceso de especiación, o cuál considerar la "unidad evolutiva especie" cuando diferentes caracteres evolucionan de a grupos de tamaños diferentes, la delimitación a veces se vuelve enteramente una cuestión de consenso.

Las gaviotas del género Larus presentan alrededor del Ártico un anillo de poblaciones cuyos extremos no son interfértiles. Los taxónomos reconocen entre 2 y 8 especies. Sobre la base de este gráfico en el primer concepto taxonómico de la taxonomía moderna Lineo hubiera nombrado 7 "tipos" en correspondencia con los 7 nombres vulgares, y hubiera reconocido la existencia de híbridos entre ellos, dejando a resolver el problema de los límites de cada especie.
Un linaje, una especie, consta de una o un grupo de poblaciones, que también son difíciles de definir, si bien el concepto espacial, sus límites geográficos, metodológicamente siempre están presentes. Normalmente se definen como un grupo de individuos de una misma especie que ocupan una región geográfica más o menos bien definida y con los individuos interactuando entre sí.[18] Estas "interacciones" no solo refieren al intercambio de genes que puede haberlo o no según su tipo de reproducción sino también al hecho de que compiten por los mismos recursos, comparten las mismas condiciones, son afectados por los mismos gremios de predadores, patógenos y parásitos y de competidores interespecíficos por algún recurso, poseen las mismas interacciones de simbiosis y comensalismo con otras especies, y se reproducen y dispersan por los mismos mecanismos, por lo que la población sufre además de los avatares del azar una selección de genes mediante la supervivencia y reproducción sexual o asexual de aquellos individuos que los porten y cuya descendencia eventualmente ocupa el nicho que ocupaban los demás.[118] (El nicho en ecología se define en función de la supervivencia del linaje[119]: las características ecológicas del género o rangos superiores no alcanzarían para explicar cómo sobreviven sus linajes; algunos taxónomos pueden utilizar una definición más amplia de nicho para referirse a los espacios dimensionales que acotan el nicho de las especies involucradas en taxones superiores, concepto que otros prefieren llamar "zonas adaptativas".[nota 9]) Estas interacciones entre individuos que mantienen integrado el linaje en un nicho diferenciado son definidas tradicionalmente como "cohesión",[cita 35] un concepto con más factores que la posibilidad de hibridar en simpatría —el "concepto de especie biológica" de Mayr—, pero que no siempre delimita unidades discretas: la cohesión tiene grados, las poblaciones se comunican entre sí por "corredores" que las mantienen más o menos cohesivas y quizás a diferente grado en cada aspecto a evolucionar, aun sin corredores los individuos de las poblaciones no se aparean al azar y se aparean o interactúan con más probabilidad con los individuos más cercanos a sí mismos, por lo que priorizar factores diferentes de la cohesión puede dar resultados diferentes. Incluso si se llega a un acuerdo en estos factores y grados, si los linajes constan de un continuum espacial que correlacione con un continuum de cohesión, todavía pueden ser divididos en unidades de forma arbitraria.[137] Esto llevaría a clasificaciones incompatibles, cuando sus delimitaciones se solapan.[117][138] En la búsqueda de la categoría se puede reconocer una cota inferior (una población que no sobrevive sin su metapoblación, es decir que no es independiente, no es una especie) y una superior (por ejemplo, la descripción de un género no debería alcanzar para explicar cómo sobrevive cada uno de sus linajes, cómo se mantiene su "continuidad" en el tiempo). Esa búsqueda al menos hoy en día parece ser infructuosa en algunos taxones, especialmente microbios de reproducción asexual, cuya cohesión parece ser un continuum, en el que todas las categorías parecen ser arbitrarias.[137]
De los conceptos de especie, que son "casi tantos como especialistas dispuestos a discutirlos", dos tuvieron importantes implicancias en la estabilidad de la nomenclatura y en las ramas de la ciencia que los utilizan.[117] Uno es el "concepto de especie biológica" o BSC (biological species concept[cita 36]), que delimita a la especie como el grupo de poblaciones que puede hibridar entre sí y dar descendencia fértil, que no subdivide las comunidades reproductivas a menos que se mantengan aisladas en simpatría, y es el concepto más utilizado, o al menos el más invocado, desde su articulación por E Mayr en los 1940-1960 hasta los 1980-1990; el otro que ganó popularidad en las últimas décadas es el "concepto de especie filogenética" o PSC (phylogenetic species concept[cita 37]), que divide las comunidades reproductivas en un grano más fino según si conservan caracteres en un análisis de filogenia, y en muchos taxones, incluso en algunos que parecían muy estables como Primates, se superpuso su clasificación a la del BSC.[117]
Una de las intenciones del BSC como formulado por E Mayr era en parte minimizar la confusión causada por la multitud de ecotipos[139] dentro de las especies biológicas.[110] Esto es porque la práctica habitual es que se describan las especies de una pequeña región dada, que en esa región están bien aisladas reproductivamente entre ellas, pero en los intentos de integrar la información entre regiones es común encontrar un continuum de tipos intermediarios.[137] Sin embargo, el BSC, fuera de los taxones de vertebrados de tamaño grande a los que Mayr estaba acostumbrado, no resultó delimitar taxones con tanta precisión como parecía, entre las especies biológicas puede haber diferentes grados en la capacidad de hibridar y quizás en un 10% de las especies biológicas de animales, un 25% de las especies biológicas de plantas y un porcentaje más alto de microbios se encuentren híbridos y no necesariamente con especies hermanas;[137] por lo que aún bajo el BSC se siguen encontrando diferencias entre taxónomos a los que se ha denominado "divisores" (splitters) y "agrupadores" (lumpers) ([117] misma terminología que en taxones superiores[140]:238-241), mientras que en los continuum sigue existiendo el problema de los solapamientos. Habiendo decidido utilizar ese concepto de especie, todavía se suman a la incertidumbre las inconsistencias históricas entre la delimitación práctica —normalmente fenética— y la mucho más costosa delimitación conceptual de las especies.[137] El requerimiento clave para solventar estas diferencias es que los taxónomos sean explícitos acerca de qué concepto taxonómico y qué método para alcanzar su delimitación están siendo utilizados, de forma de poder superponerlas y utilizarlas sincronizadamente cuando eventualmente se encuentra más de una clasificación describiendo la misma región.[117][137]
Nomenclatura biológica
El último paso en una clasificación consiste en buscar qué nombre científico le corresponde al taxón o averiguar si se necesita publicar uno nuevo, y eso se hace según las reglas escritas en los Códigos Internacionales de Nomenclatura. Por razones históricas hay uno para cada disciplina: de Zoología,[31] de Botánica,[29] y de Bacterias,[30][nota 10] y se actualizan frecuentemente como resultado de los Congresos Internacionales que reúnen a los científicos para tal efecto. Por ejemplo, a julio del 2015 el último Código de Botánica es el de Melbourne (revisión 2011, publicado en 2012[29]), el último de Zoología es la cuarta edición (revisión 1999, publicado en el 2000[31]), y el último de Bacterias es la revisión de 1990 (publicado en 1992[30]). Las ediciones pueden no ser la última versión, ya que se han publicado posteriormente enmiendas. Los Códigos poseen algunas diferencias entre sí, pero en su esencia funcionan de forma similar. Con el objetivo, y el "principio básico",[19] de la "estabilidad en la nomenclatura"[cita 38] los autores de los Códigos enumeran sus principios,[cita 39] de los que el "Principio de Prioridad" consiste en que cada taxón con su circunscripción, posición y rango debe poseer el primer nombre de los establecidos para él como único "nombre aceptado" ("nombre correcto" en Botánica, "nombre válido" en Zoología) para la comunicación.
El principio de prioridad es sencillo de expresar, lo dificultuoso es decidir cuándo dos autores se están refiriendo al mismo taxón y cuándo a uno diferente, o dicho con más rigor, lo dificultuoso es decidir, cuando un nuevo autor cambia las hipótesis taxonómicas de forma de recircunscribir los grupos o cambiarlos de posición o rango, cuál de los nuevos taxones deberá utilizar un nombre ya utilizado en el pasado. Los Códigos deciden que el nuevo taxón que "hereda" el nombre es el que contiene el mismo tipo dentro de él. Los Códigos buscan que cada descripción, cada concepto taxonómico o cada conjunto de subtaxones que ya ha sido nombrado continúe con el mismo nombre en lo posible. De esta forma los Códigos definieron lo que en nomenclatura se llama el "taxón nominal":[cita 40] la asociación permanente entre un "nombre" y un "tipo", este último establecido, en estos Códigos, en una categoría taxonómica primaria dada. Esta asociación perdura a través de los cambios de circunscripción, necesaria para establecer y reutilizar el taxón nominal pero variable entre autores; a través de los cambios de posición taxonómica, que en la categoría especie lleva a una modificación en el nombre; y a través de los cambios de categoría si se cambia a una categoría cercana que mantiene el tipo (del mismo "grupo de categorías"), que normalmente lleva también alguna modificación en el nombre.[cita 41]
Especímenes tipo En plantas, por lo general el "espécimen tipo nomenclatural" es un ejemplar completo de la especie que fue colectado y herborizado (aplastado entre papeles de diario que se cambian todos los días hasta que dejan de humedecerse), y luego depositado en un herbario accesible al público, como los de los jardines botánicos. En aves y mamíferos los especímenes tipo suelen ser animales disecados, depositados en colecciones como las de los museos de historia natural. Se conservan con más facilidad las partes duras como huesos, caparazones, valvas, exoesqueletos, y en frascos en formaldehído los animales o partes blandos. En bacterias, protozoos, algas y hongos, el tipo no suele ser un único espécimen sino una cepa cultivada viva, en general congelada o enfriada y disponible en al menos una o dos colecciones accesibles según el Código. Cada especie correctamente descripta y nombrada posee un espécimen tipo (técnicamente, es el taxón nominal el que posee un tipo). Si depositar un ejemplar como tipo es complicado, se puede reemplazar por cuidadosas ilustraciones y fotos. En los últimos años es buena práctica[99] conservar una muestra del espécimen de forma que se pueda extraer su ADN para los análisis. |


El "tipo", el contenido del taxón que se fija permanentemente al nombre, es algo diferente si estamos hablando de un nombre en la categoría especie o inferior, o de un nombre de una categoría superior a especie. En los Códigos en uso, en la categoría especie o inferior a especie (las categorías del grupo especie), "el espécimen tipo nomenclatural" es un espécimen específico de la especie idealmente completo, en buenas condiciones, y depositado en una colección cuya manutención y crecimiento sean dirigidos por un taxónomo especializado (un curador), quien debería permitir al público un acceso a los tipos aunque sea restringido. Cuando es posible, los tipos se depositan en instituciones dedicadas a la ciencia como museos de historia natural y jardines botánicos. Cuando el tipo utilizado en la publicación original (el "holotipo") se ha perdido o vuelto irreconocible, no hay otro espécimen de la colección original que pueda reemplazarlo (un "lectotipo"), y hay necesidad de un tipo para reutilizar un nombre, el nombre no se pierde sino que se puede seleccionar de los especímenes identificados con el mismo nombre y localidad que el original, un "neotipo"[cita 42]. En los casos particulares de bacterias[cita 43], protozoos[cita 44], algas y hongos[cita 45] cuyas colecciones constan de cepas vivas, el tipo no es un único organismo muerto sino que es una cepa cultivada viva, en general congelada, y disponible en al menos una[cita 43] o dos[cita 45] colecciones diferentes según el Código.

Espécimen tipo y su ubicación en la categoría de especie (en blanco los "especímenes de referencia" depositados en colecciones y no portadores de nombres) y su correspondencia con el tipo nominal (portador del nombre) en las categorías de género y familia. La distancia entre especímenes en el plano representa cuánto se diferencian en términos de sus caracteres, que suele correlacionar con la ubicación geográfica en la que fue colectado cada uno. El límite inferior de la composición de organismos de género y especie "tipo" incluidos en la familia es el mismo espécimen del taxón nominal; pero no hay ninguna obligación en los Códigos de incluir la circunscripción de la especie en el género, ni la del género en la familia, cuando ésta es establecida. Los únicos límites impuestos por los Códigos a la circunscripción del género es la dada por los tipos externos a la circunscripción de la familia que hayan sido establecidos anteriormente a la fecha del tipo aceptado; son los mismos límites impuestos para la especie en relación al género.
Con respecto a los taxones superiores a especie, el nombre del género —y de las demás categorías del grupo género— posee como "tipo" al taxón nominal de aquella especie contenida en él que fue publicada primero; y el nombre de una familia —y de las demás categorías del grupo familia— posee como "tipo" al taxón nominal del género contenido en ella que fue publicado primero. Nótese que en estos casos el "taxón nombrado" es un grupo de organismos cuyo tipo es un género o una especie contenidos en él, aunque, como está nombrado según las reglas formales de nomenclatura, el tipo contiene a su vez un espécimen tipo o un taxón que eventualmente lo anida. Con los cambios de circunscripción, ese espécimen es el único que se mantiene necesariamente dentro de los límites de cada taxón de cualquier categoría.
Si dentro de la extensión[nota 11] (la composición de organismos) del taxón no hay ningún ejemplar especificado como "tipo", el autor de la circunscripción puede decidir nombrarla formalmente, es decir, crear un taxón nominal que pasará al terreno de los "nombres científicos" por el resto de la historia de la taxonomía. El tipo será un ejemplar identificado dentro de la extensión del taxón, debidamente depositado en una colección, y el nombre será un nombre en latín o latinizado construido según ciertas reglas explicitadas en los Códigos (algunas de las cuales se verán más abajo). Todos los datos del nuevo taxón (el nombre, la posición, la categoría taxonómica, la descripción o la diagnosis escrita en forma de caracteres expresados en palabras, los datos de la ubicación del tipo) son "efectivamente publicados" si lo son donde sean accesibles para otros científicos y expertos locales, como una revista científica, o un libro. No se consideran efectivamente publicados los nombres publicados en medios efímeros de publicación como un catálogo de semillas, un diario, o por correo electrónico. Todos estos datos efectivamente publicados convierten al taxón nominal en un nombre establecido[cita 46] ("válidamente publicado" en Botánica, "disponible" en Zoología), y es cuando decimos que un nombre tiene el estatus de "nombre científico" o de "nombre formal".[cita 47]

Los dos tipos de sinonimia son los sinónimos subjetivos (Zoología), taxonómicos o heterotípicos (Botánica): los nombres que compiten por un mismo taxón, y los sinónimos objetivos (Zoología), nomenclaturales u homotípicos (Botánica), todos los taxones que "heredan" el mismo nombre, con el mismo autor original y diferente autor de la descripción (nótese que estos últimos no son verdaderos "sinónimos", son en realidad palabras ambiguas). En los casos de las especies, para las que queda explicitada la posición taxonómica (en este caso el género al que pertenecen) en el nombre, cuando una especie "hereda" el taxón nominal en un género diferente que el original, se crean dos nombres científicos con diferente género y mismo epíteto, con el mismo autor original y diferente autor de la posición taxonómica, aunque tuvieran la misma circunscripción; la ambigüedad está en el taxón nominal y solo se observa en el epíteto del nombre; el "nombre" que le corresponde a los dos taxones no es un nombre sinónimo porque son dos taxones diferentes.[cita 48]
Cuando se encuentra más de un "tipo" dentro del taxón, los demás nombres pasan a ser sinónimos taxonómicos del primero, el "nombre aceptado" ("nombre correcto" en Botánica, "nombre válido" en Zoología) del taxón. Pero si al descubrirse este hecho un nombre más reciente ya está muy extendido, puede proponerse a la Comisión respectiva como excepción al principio de prioridad y en aras de la estabilidad, agregar el nombre a la lista de nomina conservanda, nombres que se consideran el nombre aceptado por razones prácticas.
Los nombres también pueden ser establecidos pero ilegítimos. Esto es porque las reglas enunciadas para establecerlos no han impedido su formación, pero se descubre que no cumplen con los principios, los que se intentan cumplir mediante las reglas y que son enunciados en la primera parte de cada Código. Por ejemplo los principios más importantes para mantener la estabilidad en la nomenclatura son que debe haber un solo nombre para cada taxón, y el principio de prioridad. Las dos razones principales por las que un nombre puede ser ilegítimo son que al momento de la publicación no se ha evitado que (a) fuera redundante, es decir que el autor establece un nombre científico para un taxón que en la composición de organismos posee un ejemplar que ya forma parte de un taxón nominal y es así de una forma "evidente", y (b) que fuera homónimo —no sinónimo— de otro anterior, es decir que un "nombre" que ya formaba parte de un taxón nominal fuera asociado a un "tipo" nuevo. Mientras no haya un registro central de nombres incluido en las reglas estas situaciones no son evitables mediante ellas, y a pesar de que se lo intenta no son todavía fáciles de detectar, se publican miles de nombres nuevos por año[150] y solo se ha logrado establecer un registro central mandatario de nombres y publicaciones en bacterias, que existe desde 1980.[151] No se han terminado de informatizar los datos de la literatura publicada en papel para indexarlos en bases de datos digitales, y botánicos y zoólogos no consensúan la creación de "índices de nombres adoptados"[151] ("no suprimidos", dejando los nombres no indexados suprimidos por defecto) en los taxones que les tocan por temor a que unos pocos especialistas dejen publicaciones fuera de la indexación y acaparen la nomenclatura de un grupo.[138]
Cuando la aplicación estricta de un Código resulta en desestabilización de la nomenclatura, o no cumple los Principios, como se argumenta cuando se realizan propuestas de conservación de nombres o de supresión de nombres pobremente establecidos, la situación se presenta ante su Comisión respectiva, que tiene la responsabilidad de tomar una decisión al respecto.[cita 49] La composición y funciones de cada Comisión están contempladas en su respectivo Código, y tanto las propuestas como las decisiones tomadas en relación a ellas son publicadas en su respectiva revista especializada. Las decisiones tomadas por la Comisión Internacional de Nomenclatura Zoológica son publicadas en su revista, The Bulletin of Zoological Nomenclature,[152] las tomadas por los Comités auspiciados por la Asociación Internacional para la Taxonomía de Plantas (IAPT) son publicadas en su revista, Taxon,[153] y las tomadas por el Comité Internacional de Sistemática de Procariotas y Bacteriología son publicadas en su revista, el International Journal of Systematic and Evolutionary Microbiology (IJSEM).[154]
Finalmente, la nomenclatura botánica, la nomenclatura zoológica y la nomenclatura bacteriológica son independientes la una de la otra, (con pocos taxones exceptuados[nota 12]?). Como consecuencia, el mismo nombre puede ser utilizado para una planta, para un animal o para una bacteria, sin que sean formalmente nombres homónimos, y aunque no es lo aconsejable hoy en día animales y plantas pueden estar compartiendo el 13% de sus nombres en la categoría de género.[155]
Categoría taxonómica
|
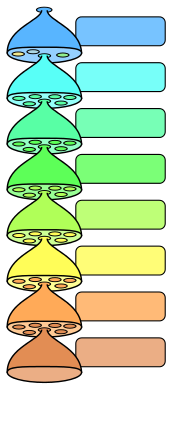
En los Códigos de Nomenclatura, son siete las categorías primarias o principales:[cita 50] Reino, Filo o División, Clase, Orden, Familia, Género, Especie. Históricamente, el Código de Zoología utilizó la categoría Filo (Phylum en inglés y en latín), y el Código de Botánica y el de Bacterias escindido de él, la de División (División, en inglés, Divisio en latín), hoy en día en una dirección hacia la armonización se los trata como sinónimos.[cita 50]
Una regla mnemotécnica para recordarlas es la siguiente:
| El rey es un filósofo de mucha clase que ordena para su familia géneros de buena especie | ||
- Reino Filum Clase Orden Familia Género Especie | ||
| La reina es una diva de mucha clase que ordena para su familia géneros de buena especie | ||
- Reino División Clase Orden Familia Género Especie | ||
El establecimiento de un nombre en una categoría primaria genera un "tipo" en lo que los Códigos llaman el "grupo de categorías" que abarca todas las categorías intermedias por arriba y por debajo de la categoría primaria, que poseerán, justamente, el mismo "tipo". Estos taxones serán establecidos cuando se los publique por primera vez, con su respectiva circunscripción.[cita 50]
Luego de establecer las categorías primarias pueden intercalarse algunas categorías secundarias, pero a diferencia de las primarias estas son optativas, y no es necesario intercalarlas a lo largo de toda la categoría en una clasificación, pero una vez que se decidió agregar una categoría secundaria a un taxón primario, sí es necesario agregarla a lo largo de todo ese taxón. Son tribu entre familia y género (perteneciente al grupo familia), sección entre género y especie (perteneciente al grupo género), variedad y forma debajo de especie, y las que utilizan prefijos: superfamilia, subfamilia.[cita 50] La categoría de subtribu sólo puede establecerse después de la de tribu, y la de subsección después de la sección. Las categorías secundarias propuestas por cada Código se presentarán en la sección siguiente.

Las categorías secundarias se intercalan de forma de mantener los conceptos taxonómicos en el rango que les corresponde, en la opinión de un autor.
Nombre científico
En los Códigos de Nomenclatura, cada especie queda designada por un binomio (una expresión de dos palabras) en latín, donde la primera palabra, el "nombre de género", es compartida por las especies del mismo género; y la segunda, el "adjetivo específico" o "epíteto específico", hace alusión a alguna característica o propiedad distintiva de esa especie en particular, como pueden ser el color (albus, ‘blanco’; cardinalis, ‘rojo cardenal’; viridis, ‘verde’; luteus, ‘amarillo’; purpureus, ‘púrpura’; etc.), el origen (africanus, ‘africano’; americanus, ‘americano’; alpinus, ‘alpino’; arabicus, ‘arábigo’; ibericus, ‘ibérico’; etc.), al hábitat (arenarius, ‘que crece en la arena’; campestris, ‘de los campos’; fluviatilis, ‘de los ríos’; etc.), un homenaje a una personalidad de la ciencia o de la política o atender a cualquier otro criterio. No es necesario que el nombre esté en latín, sólo es necesario que esté latinizado. Los nombres de género siempre van con la primera letra en mayúsculas, los epítetos específicos siempre van en minúsculas, y los nombres de género y de especie van siempre en itálicas (o subrayados, si se escribe a mano).[nota 13] Al escribir el nombre de especie, el epíteto específico nunca es utilizado solo, y es mandatario que esté precedido por el nombre del género, de forma que el nombre de la especie sea el binomio completo. La abreviación a la primera letra del nombre del género precediendo el epíteto específico completo también es aceptable una vez que el nombre ya apareció en su forma completa en la misma página o en un artículo pequeño. Así por ejemplo, la lombriz de tierra fue llamada Lumbricus terrestris por Linneo, y si el nombre ya apareció antes en el artículo y no hay ambigüedad, puede volver a referirse a ella como L. terrestris. Con respecto a los taxones ubicados en la categoría de género y superior, los nombres son uninomiales (constan de una sola palabra) y siempre se escriben con la primera letra en mayúsculas (aunque solamente en la categoría de género van en itálicas[nota 14]). El epíteto específico debe concordar gramaticalmente con el género, y si en su género original no concordaba se modifica para que lo haga: Leucadendron argenteum fue originalmente descripto en otro género como Protea argentea. Como los Códigos de Nomenclatura prohíben que dentro de cada Código haya dos taxones con el mismo nombre, es decir dos taxones nominales con el mismo nombre, no puede haber dos géneros con el mismo nombre (ni dos taxones por arriba de género con el mismo nombre), pero como ocurre que el epíteto específico de las especies solo se usa después del nombre del género, puede haber dos especies diferentes pertenecientes a géneros diferentes que compartan el mismo epíteto específico. Una vez fijado, un nombre no es sustituido por otro sin un motivo nomenclatural. Por ejemplo el roble de los alrededores de Madrid fue bautizado como Quercus pyrenaica erróneamente, puesto que no se encuentra en los Pirineos, pero tal circunstancia no justifica un cambio de nombre.
La recomendación es que los nombres sean eufónicos, fáciles de memorizar y descriptivos,[cita 53] y a que los lectores aprendan a obtener ese atractivo con el objetivo de que sean utilizados por una parte significativa de los usuarios finales dedicó Linneo muchas de sus secciones en Philosophia Botanica (1751[139]); pero no hay reglas que impidan a los investigadores utilizar su propia inventiva para crear los nombres científicos más insólitos: el nombre de la araña Pachygnatha zappa, porque tiene una mancha en el abdomen igual al bigote del artista Frank Zappa; algunas moscas chupadoras de sangre del género Maruina: Maruina amada, M. amadora, M. cholita, M. muchacha, M. querida, M. chamaca, M. chamaguita, M. chica, M. dama, M. nina, M. tica y M. vidamia, todos adjetivos cariñosos; el dinosaurio que fue llamado Bambiraptor debido a Bambi, el de la película de Disney; el molusco bivalvo Abra cadabra (aunque después se lo cambió de género); y quizás el caso más sobresaliente, el género de arañas brasileñas Losdolobus, llamado así porque los investigadores que lo describieron, queriendo homenajear a dos argentinos que habían colaborado en parte significativa en la publicación de la que no iban a ser los autores, les pidieron a estos que inventasen un nombre para el nuevo género, que quedó como Losdolobus por "los dolobus", término del lunfardo argentino intraducible en una enciclopedia.[158] Los Códigos en una de sus recomendaciones hacen un llamado a la responsabilidad, "Los autores deberían ejercitar un razonable cuidado y consideración en la formación de nombres nuevos para asegurarse de que son elegidos con sus subsecuentes usuarios en mente y que, mientras sea posible, sean apropiados, compactos, eufónicos, fáciles de memorizar y no causen ofensa".[cita 53]
En las categorías superiores a género los nombres científicos están compuestos por la raíz del tipo (un género del taxón, en los rangos más bajos el más antiguo) seguida de un sufijo que denota la categoría en la que el taxón fue ubicado,[cita 54] como se indica en la siguiente tabla:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Un "nombre descriptivo" es por ejemplo Monocotyledoneae ("un cotiledón"), o un nombre con una raíz descriptiva en lugar de derivar de un género tipo como Coniferales (raíz: "Conifer-" sufijo: "ales").[cita 54] Estos nombres no están asociados a un género "tipo" en Botánica, sino a la descripción inferida del nombre (p.ej. las monocotiledóneas pueden recircunscribirse de forma de dejar cualquiera de sus subtaxones fuera del grupo, pero poseer "un cotiledón"), en cambio sí están asociados a un género tipo en Bacterias, además de a la descripción inferida del nombre[cita 54]. Los Códigos llaman a los nombres descriptivos sin género tipo typeless ("sin tipo"), algunos autores prefieren definir como "tipo" al contenido del taxón que esté asociado al nombre en un taxón nominal, por lo que la descripción inferida de un nombre descriptivo sería también un "tipo". Nótese que esta regla difiere de la de taxones inferiores, en los que la descripción inferida del nombre descriptivo nunca se agrega al "tipo". Además en taxones superiores al grupo familia, o superiores al grupo orden en bacterias, no es mandatario que el género seleccionado como "tipo" sea el primero de los nombrados en regla.
Un ejemplo de taxón es el orden Primates. En esta expresión, "orden" específica la categoría taxonómica del grupo, más abarcativa que la categoría de familia y menos abarcativa que la de clase. "Primates" es el nombre del taxón. El orden Primates está incluido en la clase Mammalia (los mamíferos), y a su vez incluye diversas familias como la familia Cebidae (cébidos, las monas americanas) o la familia Hominidae (homínidos, la familia que nos incluye como especie).
Por debajo de la categoría especie se encuentran la subespecie, variedad y forma, todas categorías pertenecientes al "grupo especie". Su nombre se construye empezando con el nombre de la especie seguido de espacio y el conector subsp. o var. o f. respectivamente (sin itálicas), seguido de un epíteto subespecífico nuevamente en itálicas: Carpinus caroliniana subsp. caroliniana, Lyonia ligustrina var. ligustrina, salvo en las subespecies de animales que no llevan la partícula subsp: Canis lupus familiaris. El epíteto subespecífico debe concordar gramaticalmente con la especie, y si en su taxón nominal no concuerda se modifica para que lo haga. Son nombres trinomiales.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Además, en todos los taxones, la primera vez que se nombra un taxón en una publicación científica, el nombre científico puede verse seguido del apellido del autor de su primera descripción, que es el que estableció el taxón nominal (normalmente llamado la "autoridad"), mejor aún si se sigue del año en que el taxón fue establecido. El nombre del autor puede estar abreviado, por ejemplo a Linneo se lo abrevia con una «L.». Las veces posteriores que se nombra al taxón en la publicación se evita el autor y el año de publicación. A veces se encuentran variaciones en la forma en que autor y año son escritos (por ejemplo hay dos autores conectados por palabras en latín como in o et o ex, o el autor está entre paréntesis), y cada una de estas variaciones tiene su significado en Nomenclatura. Por ejemplo, en el caso de los animales, si en la actualidad una especie en particular está ubicada en un género diferente al que le fue asignado por su autoridad original, el nombre del autor y el año se ponen entre paréntesis indicando así que originalmente se la describió con el mismo taxón nominal, pero otro nombre. Así, el león en las clasificaciones actuales es «Panthera leo (Linnaeus, 1758)», y se pone autor y año entre paréntesis porque Linneo lo describió originalmente como «Felis leo» (Lineo seguía el nombre del taxón del autor de su concepto taxonómico, que no siempre coincidía con el primer autor del nombre[139]). El autor y año de publicación no son reglas sino recomendaciones de los Códigos de Nomenclatura, pero son de uso corriente y se encuentra en la literatura taxonómica que son altamente recomendados por muchos autores, probablemente debido a que no hay registro central, lo que vuelve las publicaciones taxonómicas muy tediosas de encontrar, quien quiera encontrar una en particular debe recorrer las hemerotecas y bibliotecas de todas las Universidades, Museos de Historia Natural y Jardines Botánicos y aun así nunca estará seguro de que una publicación no exista porque no la haya encontrado. Las publicaciones taxonómicas no están clasificadas como tales en las revistas que las publican ni en las hemerotecas que las reciben, ni hay acceso público a una "lista de taxónomos" en carrera. Cuando se hace referencia a la circunscripción de un concepto taxonómico utilizado en la actualidad que difiere de la circunscripción de la publicación del taxón nominal, se puede encontrar el conector sensu,[159] p.ej. «Arenaria sensu McNeill (1962)», o como en los casos en que se diferencia sensu lato (cuya circunscripción involucra el mayor número de organismos, no necesariamente de subtaxones) del mismo nombre sensu stricto (cuya circunscripción involucra el menor número de organismos). El autor y año del taxón nominal original se mantienen útiles para evitar errores con nombres homónimos. Esperablemente, en el futuro esta notación pueda reemplazarse de forma de desambiguar todos los conceptos taxonómicos asociados al mismo nombre en un registro central de nombres. En informática de la biodiversidad, se propone utilizar un Identificador Global de Conceptos Taxonómicos (de nombres según cada autor), que al resolverse guíe a la cita del concepto utilizado con todas sus referencias y quizás un enlace a la publicación y a los usos previos del nombre hasta el momento en que fue establecido.[34][160][159] La idea de hacer pública a la Ciencia es permitir una evaluación que no permita que se pierdan los estándares más simples.
¿Por qué cambian los nombres de los taxones?
La descripción formal y la tipificación de los taxones son procedimientos en los que es frecuente observar errores e inconsistencias, y esto ocurría aun en mayor medida antes de que los Códigos Internacionales reglamentaran estos trabajos muy pormenorizadamente. Los nombres de los taxones pueden cambiar cuando son redescubiertas descripciones más antiguas del mismo taxón, y se aplica el principio de prioridad, o cuando se observa que la descripción original en realidad refería a un taxón diferente. No suelen darse estos casos en los grupos ya bien estudiados, y en los Códigos hay además mecanismos para la supresión de nombres pobremente definidos en los grupos más complicados. Una vez asentada la taxonomía fundamental de un grupo, es poco probable que los cambios de nombres se den por esta clase de motivos.
Los nombres cambian cuando el descubrimiento de nueva información obliga a modificar las circunscripciones, pero también —y sin necesariamente nueva información que lo justifique—, los nombres cambian por diferencias de criterio entre los especialistas.
Por ejemplo, es muy común que a distintos grupos de organismos recién descubiertos se les dé nombres de especie (y de género incluso) nuevos y que años después un taxónomo que los estudie llegue a la conclusión de que todos pueden hibridar entre sí y dar descendencia fértil, y unifique todos los taxones en una única especie (en este caso todos los nombres cambian al nombre de la primera especie descripta dentro del grupo), dejando, en todo caso, los antiguos epítetos de especies como subespecies. Con posterioridad puede llegar otro taxónomo que considere que esos grupos de organismos deberían dividirse en dos especies distintas, que en la naturaleza conviven sin hibridarse (aunque pudieran), debido a que cada una de las especies que él propone forma una unidad monofilética en un análisis de filogenia. Entonces a los que se encuadran en la "nueva" especie, se les cambia el nombre de especie por el nombre de la especie más antigua descripta dentro de la "nueva" especie. Otro taxónomo podría argumentar más tarde que todos los grupos originalmente descritos son especies válidas del mismo género, debido a que cada uno de ellos puede ser identificado por su propio grupo de caracteres morfológicos, y que dentro de cada uno de ellos hay un patrón de ascendencia y descendientes. En este caso, todos los epítetos de especies cambiarían al primer epíteto dado a cada grupo cuando fue descrito, pero todos los nombres de género se unificarían en uno solo, por lo que todos los géneros pasarían a nombrarse como el género más antiguo descrito dentro del grupo. Esta inestabilidad de los nombres se habría producido casi únicamente por cambios de "filosofía taxonómica": el "concepto taxonómico" que cada autor considera que se corresponde con la categoría especie. Estas diferencias conceptuales entre taxónomos son extremadamente comunes hoy en día, lo cual lleva a una gran inestabilidad potencial en los nombres de los taxones.[161]
Determinación o identificación de especímenes

Captura de insectos voladores en el estrato herbáceo. La primera actividad de un aspirante a conocer la taxonomía de su región es crear su propia colección, producto de colectas periódicas a lo largo de todas las estaciones del año, clasificada hasta que logra identificar informalmente los cientos o pocos miles de especies o morfotipos nativos y adventicios presentes en su localidad, y establecer las "diferencias" entre los más cercanos hasta conceptualizar los caracteres o los atributos que los delimitan.[139] El paso siguiente es la comparación con los nombres formales, con ayuda de un taxónomo especializado o con la bibliografía taxonómica recomendada por él.[139]

Caja entomológica de dípteros, contiene una plancha de telgopor en la que se insertan los alfileres entomológicos en los que están montados los dípteros y sus etiquetas. El aspirante debe tener conocidos todos los grupos "naturales" por arriba de género o de familia (en este caso orden Diptera, moscas y mosquitos) para realizar este paso al segregar los especímenes de su colección.[139]
La disciplina de la Determinación, o identificación de especímenes, es el proceso de tomar los atributos de un espécimen hasta encontrar el taxón en el que se ubica ("determinarlo") o reconocer que es nuevo para la ciencia y necesita una descripción formal y un nombre. Como sólo el 10 % de las especies del planeta está descripto, y esas descripciones son hipótesis con diferentes grados de resolución,[cita 9][162] la Determinación funciona recursivamente con la descripción y delimitación de especies en el área llamada alfa-Taxonomía o Taxonomía Descriptiva, o, modernamente y deslindando las diferencias en las definiciones de Taxonomía —desde hace décadas con una definición minimalista— y Sistemática, y debido a que se integradan (se van indiferenciando a medida que el foco de atención se traslada de una a la otra) en esa categoría taxonómica, el área de la "Sistemática Descriptiva"[23] o "Ciencia de la Biodiversidad"[cita 7]. Esta área se ocupa además de la investigación expedicionaria y exploratoria en ambientes naturales para avanzar en el descubrimiento, la identificación, la descripción, la clasificación y la catalogación de la biodiversidad del planeta, utilizando como unidad de trabajo el organismo y cuya prioridad[24] es la descripción de las especies dentro de un contexto evolutivo, aproximándose a ello desde la investigación primaria sobre un grupo taxonómico dado, un gremio ecológico, o una región geográfica;[cita 7] se ocupa de realizar nuevas colectas (el "trabajo de campo") y depositarlas en colecciones, del trabajo de toma de datos sobre las colecciones existentes, de la realización de clasificaciones revisionarias, de desarrollar hipótesis taxonómicas (determinación de homologías, caracteres, apomorfías y plesiomorfías, especies, clados[24]), de testear las hipótesis taxonómicas de especie, y la creación y uso de las herramientas informáticas necesarias para todo ello y para publicar las nuevas claves de identificación y las nuevas descripciones, con las que en conjunto se llega a los taxones redefinidos, que junto con el desarrollo teórico de las hipótesis forman parte de la literatura taxonómica primaria, cuyos productos más acabados[23] son las llamadas floras, faunas y monografías taxonómicas.[cita 7] Durante una determinación, todos los puntos tomados de la literatura deben ser comprobados con los especímenes propios para tomarlos por verdaderos.[139] Especialmente en áreas donde la biodiversidad presenta una tarea desafiante, como en los trópicos –debido al número de especies y también al vasto número de especies que no ha sido aún publicado, o exhaustivamente fotografiado– el rol del especialista, profesional o experto local, en la determinación es crítico.[18]:7
Además de la finalidad en sí misma de generar una taxonomía sólida y estable, hay varias otras razones por las que se puede necesitar una determinación, y todas si están correctamente documentadas y los ejemplares depositados en colecciones con acceso al público general y a la comunidad científica, aportan datos a la alfa-Taxonomía. Los taxónomos necesitan identificar los organismos sobre los que extraen nuevas líneas de evidencia con las que ajustar los análisis de filogenia en taxones de categorías más altas, y a pesar de que se elige para sus muestreos a los especímenes que se consideran bien determinados tampoco ha faltado algún error en la reconstrucción filogenética por un error de arrastre de determinación.[164] En Biogeografía se utilizan los datos de especímenes identificados por localidad u "ocurrencias", lo que se llama "datos georreferenciados", en los análisis,[165] cuyas conclusiones a su vez pueden debilitar o robustecer hipótesis taxonómicas. Muchos trabajos de investigación realizados por biólogos ecólogos implican un conteo de alguna unidad taxonómica de biodiversidad en cierta área, como en las investigaciones relacionadas con la conservación,[166] o en las relacionadas con las invasiones biológicas y su impacto en el ecosistema, o en los cambios en patrones de sucesión en relación a algún otro tipo de disturbio. Una correcta identificación se necesita cuando se sospecha que una enfermedad o muerte en humanos, animales o plantas es causada por un ser vivo, como cuando se sospecha que el ganado está consumiendo una planta tóxica que le produce enfermedad o muerte,[167][168] o cuando un animal salvaje pica o muerde a un humano, o un microbio patógeno lo infecta,[169] la correcta identificación en estos casos abre la literatura que indica las formas de control y tratamiento conocidas, y estas ocurrencias agregadas pueden analizarse en la búsqueda de correlaciones y patrones predictivos. Los fabricantes de pesticidas y herbicidas[170] y los servicios relacionados con la salud pública[171] pueden requerir un monitoreo de insectos o plantas dañinos en cada zona con el objetivo de mantener regulada su abundancia y para probar efectividad de tratamientos cuando se decide aplicarlos. En el campo de la biología forense la identificación de especímenes esclareció asuntos legales.[172][173]

Ejemplo de Argentina. Total de ocurrencias (todos los taxones, todas las épocas) publicadas en GBIF (izquierda) y en BOLD (repositorio de la línea de evidencia ADN "barcode" COI, derecha), a septiembre del 2016. GBIF: 979 datasets totalizando algo más de un 1,8 millón de récords (ocurrencias).[174] BOLD: Casi 100 mil récords (ocurrencias) de los que algo más del 5% está determinado hasta especie.[175]
Si se han realizado identificaciones para un trabajo que se está por publicar, es buena práctica[176][177] citar cuál fue el taxónomo consultado o las publicaciones taxonómicas que fueron utilizadas para identificar las especies, y agregar una sección de Especímenes examinados[178] donde se cite la ubicación de los especímenes que fueron depositados o visitados en la colección que los preserva como referencia (los voucher[179]) de los datos sobre los que se realizó la hipótesis publicada. La sección "especímenes examinados" debería incluir el nombre científico actual [o el aceptado sensu qué autor], el número de identificación del espécimen individual dentro de la colección (el accesion), el nombre de la colección o su acrónimo, la localidad donde fue colectado, y si se las obtuvieron, los accession de las secuencias de ADN publicadas por ejemplo en GenBank.[178] Si fue publicado el dataset de ocurrencias por ejemplo en GBIF, puede citarse la ubicación del mismo en el repositorio público en el que fue compartido o las ocurrencias dentro del mismo utilizadas. Las colecciones de especímenes poseen una infraestructura específica para cada taxón, y cada autor puede tener una colección privada —incluso unicata (unicate collection[156]), con un solo espécimen o lote, quizás holotipo o isotipo—, pero si es posible se alojan[180] en instituciones públicas como museos de historia natural o jardines botánicos, que son los sitios más estables y las hacen perdurar por centurias, el taxónomo especializado que dirige su manutención y crecimiento –el curador– idealmente debería permitir al público un acceso a los especímenes aunque sea restringido. Las colecciones suelen tener catálogos electrónicos con la información de los especímenes que poseen, que son cada vez más fáciles de acceder y, a pesar de que en sus inicios carecían de estandarización en sus formatos, se prevé que en el futuro muchas de esas bases de datos puedan ser accedidas por un mismo portal, esperablemente mediante los estándares TDWG. Uno de esos portales, creado en el 2001 por acuerdos internacionales entre agencias gubernamentales después de la Convention on Biological Diversity (CBD) es GBIF, que aloja tanto datasets compartidos desde los catálogos de museos y herbarios como datasets que se publican asociados a publicaciones científicas. En las colecciones, no es raro que los especímenes se reclasifiquen si están lo suficientemente bien documentados, se calcula que un porcentaje que puede llegar a ser de hasta un 20% o un 25% de los especímenes puede ser reclasificado durante el rechequeo (ver[176]); algunos de ellos podrán ser designados como el "tipo"[180] de un nombre de especie nueva o de una ya nombrada cuyo tipo original se ha perdido[18].[20][181][182] Son cada vez más comunes los softwares que formatean la sección de especímenes examinados a partir de la planilla de datos, y también puede formatearse en forma de data paper un set de ocurrencias para publicación formal.
La colecta
Sólo si las circunstancias lo permiten, porque pueden ser difíciles de colectar (p.ej. pueden honguearse muy rápido...) o estar en peligro de extinción -y esta información debe obtenerse a priori en la región a trabajar-, los especímenes a identificar pueden colectarse con cuidado de no dañar ninguno de sus caracteres. No se debe perder el dato de la localidad de la que se extrajeron ni el momento en que fueron colectados, un "espécimen/localidad-momento" es una ocurrencia, las ocurrencias pueden estar respaldadas por colectas, por fotografías o ser sólo el ingreso de un dato, si la confiabilidad del colector lo permite. Para cada espécimen colectado o fotografiado se realiza una ficha de colección en la que se anota localidad, fecha y colector; y otras observaciones que pueden perderse al llegar al lugar de identificación: hábitat, su abundancia en esa localidad, si estaba asociado a alguna otra especie, si era de noche o llovía, el método por el que fue colectado, color y textura de los órganos —que podría cambiar durante la preservación—, fragancia, los id de las fotografías asociadas, etcétera ([18]:553). Las fichas de colección suelen ser planillas estandarizadas, de esta forma es más difícil olvidar anotar un dato. Si se colecta más de un espécimen, lo más prolijo es hacer una ficha de colección para cada ejemplar. Luego de la identificación, debe agregarse la misma a la ficha y también quién la realizó —ya que no necesariamente es la misma persona que realizó la colecta—, o cuáles fueron los métodos utilizados para la identificación (claves o colecciones con las que se lo comparó), para dejar lo más explícita que se pueda la circunscripción utilizada para ese nombre aceptado. Durante la campaña se numeran los especímenes y después de la misma se asocian los números a un código que haga la campaña única, que concatenados funcionan como un id (identificador) para cada ocurrencia. Finalizada la campaña, se posee típicamente un espécimen, una ficha de colección y un set de fotos asociados a cada id, los que hayan sido colectados también tendrán un número de accession en una colección (según la codificación de la colección a la que hayan sido enviados, puede no ser el mismo que el id del espécimen en la colecta, y no necesariamente cada espécimen tiene un solo número de accesion, a veces un solo accesion alberga un lote de especímenes), y esperablemente también posee asociada una lista de los estados de sus caracteres, que fueron convenientemente dejados por escrito en alguna planilla estandarizada durante la identificación. Cada id de espécimen representa una fila en una colección de "ocurrencias". Si se ingresan las fichas en un formato electrónico, debe preverse que cumpla con los estándares provistos por el Biodiversity Information Standards (anteriormente TDWG) para facilitar su acceso (ver el estándar para ocurrencias Darwin Core).
La identificación (Determinación)
El primer paso en una identificación es una observación a simple vista, que implica una apreciación inconsciente de muchos caracteres del espécimen al mismo tiempo, en la que se asocia el espécimen a cierto taxón formal o informal, filogenético o no (lo que se llama apreciación gestalt u holística del espécimen),[183] los métodos más formales de identificación entran en juego cuando el interesado necesita que la identificación continúe hasta taxones más específicos, o cuando necesita comparar sus morfotipos con las especies publicadas.([18]:6-7;[183])
Los taxónomos concuerdan en que la mejor forma ([20]:498) y la más rápida ([18]:6-7) de identificar un espécimen con métodos formales es mostrárselo al especialista, taxónomo o experto local, al que le sean familiares las especies de ese taxón superior en esa región, filogenético o no (puede haber especialistas en las plantas vasculares de una región dada, que se especialicen más en una familia de plantas en todo el planeta, o en todos los taxones de "plantas suculentas" de la región o el globo), que tenga comprobada la existencia de las especies locales en su propia colección (Lineo 1751[139]) y esté al tanto de las últimas actualizaciones taxonómicas publicadas, a las que tiene acceso en la biblioteca taxonómica local o regional. El especialista en muchos casos podrá identificar la especie casi a simple vista, y si los necesita puede faciliar los instrumentos y las herramientas literarias necesarias para identificar los especímenes que faltan de la colección.
Los instrumentos para la identificación varían según el taxón superior tratado. Para plantas y artrópodos, los instrumentos son una lupa de disección, los elementos para manipular la muestra en la lupa (dos agujas de disección y una pinza de punta fina), en el caso de las plantas un bisturí para seccionar los órganos a observar en la lupa (ovario, pimpollo, pecíolo...), puede ser útil un calibre ([18]:557) para hacer las mediciones, y a veces un microscopio. Si además se puede fotografiar el espécimen vivo o fresco la tarea de identificación se ve facilitada. Raramente dos especies no pueden discernirse morfológicamente y se debe recurrir al envío de muestras para la extracción y análisis de ADN.
- Herramientas para la identificación («determinación») de plantas y artrópodos

La lupa de los laboratorios se llama lupa de disección, permite tener las manos libres para manipulación, tiene dos rueditas para calibración gruesa y fina de la distancia al objeto, y dos lentes sucesivos, uno fijo y uno desmontable y que se vende de diferentes aumentos (si la lupa es binocular, uno desmontable para cada ojo), que logran en conjunto un aumento de hasta 40x. En la foto una lupa binocular. En inglés las lupas de disección están clasificadas como un tipo de microscopio.
Homemade dissecting needle and a flora.jpg
Aguja de disección para manipulación bajo la lupa (normalmente como en la foto fabricadas a mano, insertando una aguja de coser firme en el mango de una birome), y una flora detrás.

Pinza de punta fina para manipulación bajo la lupa.

Bisturí de hoja delgada (mango de bisturí n° 3) para seccionar órganos de plantas, como el ovario (para ver placentación), el pimpollo (para ver prefloración), o un pecíolo (para ver la estela, la disposición de haces vasculares en el corte transversal). Si se consigue, puede ser reemplazado por una hoja de afeitar.

El microscopio óptico tiene un rango de aumento de 40x a 400x. La diferencia con la lupa es que la muestra se observa "montada en un portaobjetos" (presionada entre dos placas de vidrio, portaobjetos y cubreobjetos, hasta formar una capa delgada). La muestra se observa por transparencia, por luz transmitida. El microscopio puede usarse para ver la morfología de los esporangios de los helechos, que sólo deben espolvorearse sobre el portaobjetos y cubrirse con el cubreobjetos, pero muchas veces para ver una muestra es necesario cortarla en secciones delgadas, idealmente con un micrótomo, y a veces también teñirla, este trabajo entra en el campo de la histología.

Las lupas de mano o de bolsillo, que se pueden llevar al campo, tienen un aumento de unos 2x a 6x. Son útiles para descartar en el campo especies de las que ya se tengan muchos ejemplares y llevar colectadas sólo las especies del mismo género que sean más raras o todavía no estén representadas en la colección. Las lupas con tapa rebatible son similares a la de la foto.
Field guides and a camera.JPG
Guías de campo, útiles para conocer a priori algunas de las especies o morfotipos a colectar. Cámara de fotos para fotografiar los especímenes frescos, que después de colectados pueden ser más difíciles de identificar.

Calibre. ([18]:557)





Las herramientas literarias que puede facilitar el especialista incluyen claves de identificación, que tienen en cuenta pocos caracteres, expresados además de forma "tipológica"[184], resultando ser una simplificación de la apreciación gestalt del taxónomo que las realiza; pero siempre que se dispone de ellas, es importante seguirlas. Comparar directamente con las descripciones o las fotos puede inducir errores, ya que en las fotos pueden no mostrarse o no ser evidentes para el lector no entrenado los caracteres que diferencien el taxón de los demás.[170] Las claves de identificación piden tomar los estados de los caracteres del espécimen de una forma ordenada que descarta posibilidades hasta llegar al nombre del taxón. De ellas el tipo de clave más difundido en la actualidad ([18]:557) es la clave dicotómica.
En la clave dicotómica se elige siempre uno de dos caminos, dos descripciones mutuamente excluyentes de los estados de algunos caracteres. La elección envía al usuario al siguiente par de descripciones, hasta que, si el usuario no cometió un error, se llega al nombre del taxón ([18]:558,[20]:496-497).
Clave dicotómica de géneros de la familia de las ninfáceas (una familia de nenúfares) (tomada de la Wikipedia):
- Flores hipóginas. Ápice del gineceo formando un disco plano a ligeramente cóncavo con estigmas radiales. Sépalos 5-14. Polen anasulcado, espinoso.
- Subfamilia Nupharoideae Mot. Ito, 1987.
- Género único: Nuphar Sm. in Sibth. & Sm., 1809. Holártico.
- Flores períginas a epíginas. Ápice del gineceo en forma de copa, toda su superficie estigmática. Sépalos 4-5. Polen de otro tipo.
- Subfamilia Nymphaeoideae Arn., 1832.
- Plantas con acúleos. Hojas escutiformes, peltadas.
- Borde del limbo levantado formando un reborde continuo. Envés foliar aculeado. Estaminodios y apéndices carpelares presentes.
- Género Victoria Lindl., 1837. América meridional tropical.
- Borde del limbo no levantado. Haz y envés foliares aculeados. Estaminodios y apéndices carpelares ausentes o inconspicuos.
- Género Euryale Salisb., 1805. Asia, de la India al Japón.
- Plantas sin acúleos. Hojas de diversas formas, peciolo inserto en una escotadura del borde del limbo.
- Sépalos 4-5. Estambres péndulos, epipétalos sobre la cara interna del tubo de la corola. Semillas espinosas, sin arilo.
- Género Barclaya Wall., 1827. Sureste de Asia.
- Sépalos 4(-5). Estambres erectos, períginos sobre la cara externa del tubo de la corola. Semillas lisas, papilosas o pelosas, con arilo.
- Pétalos 0-5. Eje floral central prominente por encima de la copa estigmática. Hojas sumergidas lineares con bordes fuertemente ondulados.
- Género Ondinea Hartog, 1970. Australia occidental.
- Pétalos (6-)8-40(-50). Eje floral central visible como pequeña proyección en la base de la copa estigmática. Hojas sumergidas (si existen) anchas, de bordes apenas ondulados.
- Género Nymphaea L., 1753. Cosmopolita.
La clave se llama politómica si se dan más de dos opciones en cada bifurcación, es una clave politómica en dos pasos la utilizada por Linneo en Species Plantarum (1753). En el primer paso pedía contar el número de estambres, y en el segundo el número de estigmas.
En la clave multi-acceso o policlave, se puede utilizar cualquier conjunto de caracteres en cualquier orden, es el usuario quien selecciona los caracteres utilizados para la identificación. Las claves digitales normalmente son de tipo multi-acceso, se ingresan los datos que se pueden tomar y el programa devuelve los posibles taxones a los que pertenece el espécimen. Estas claves normalmente contienen mucha más cantidad de caracteres que las claves dicotómicas. Un ejemplo es el sistema DELTA ("Descriptive Language for Taxonomy") (Askevold y O'Brien 1994,[185] Dallwitz 1993,[186] 2000,[187] Dallwitz et al. 2002,[188] Pankhurst 1978,[189] 1991,[190] Watson y Dallwitz 1991,[191] 1993[192] citados en Judd et al. 2007[18]) desarrollado en los '1990, en la web se pueden encontrar otros como el sistema LucID (Norton 2012[193]), y se espera que se vuelvan más importantes en el futuro[194]. Los programas preexistentes sirven tanto para utilizar claves ya hechas con esos programas como para construirlas a partir de una matriz de taxones × caracteres —también son relativamente fáciles de programar si se tienen nociones de programación—, y están adaptados a que se agreguen fotos, videos y sonido. Hoy en día, la mayor desventaja de las claves multi-acceso es su disponibilidad ([18]:558,[20]:496-497). Algunos ejemplos: las que identifican las plantas de cualquier región del mundo hasta familia (como en colby,[195] discoverlife[196]), hormigas (antweb[197]), saltamontes (Common Western Grasshoppers[198]), mariposas y polillas (Butterflies and Moths[199]), y muchas todavía están siendo creadas (Farr 2006[200]). Sin un registro central y una etiqueta explícita de que una clave entra en el campo de la literatura taxonómica primaria y los taxones que abarca, son difíciles de encontrar, y nuevamente debe recurrirse al taxónomo especializado o a la hemeroteca y biblioteca de las Universidades, Museos de Historia Natural o Jardines Botánicos que abarcan la región.
Si bien las claves informáticas son valiosas, la mayor parte de las claves de identificación todavía se encuentra, junto con las respectivas descripciones e ilustraciones de cada taxón, en enormes volúmenes en papel publicados en series llamadas Floras y Faunas, que reúnen las descripciones y claves de las plantas y animales respectivamente, que pertenecen a un área particular, a una pequeña región, o a todo un continente ([18]:560). La primera Flora de la taxonomía moderna, Species Plantarum (1753) de Lineo, reunía la información global hasta el rango de especie y variedad en los conceptos taxonómicos del autor, totalizando unas 8000 especies; en la actualidad la flora global editada por Kubitzki fue iniciada en 1990, llega hasta el rango de género, y todavía no está terminada —los volúmenes se siguen publicando esporádicamente 25 años después—. Las Floras y Faunas son esfuerzos conjuntos en los que un taxónomo editor le da a cada taxónomo o pequeño grupo de ellos a describir exhaustivamente el taxón para el que es especialista, en general hasta la categoría de especie en una región dada, convenientemente revisado, chequeado con las colecciones existentes y la bibliografía previa convenientemente revisionada; en general estos proyectos tardan décadas en completarse y pueden quedar incompletos si los subsidios en general gubernamentales dejan de renovarse, o si se pierden los incentivos para terminarlos. Un problema que deben enfrentar los usuarios de estas claves es su antigüedad, muchas veces su taxonomía está desactualizada. Los nombres recién descriptos para la ciencia pueden no encontrarse en ellas, ni las nuevas revisiones de géneros y familias con especies exóticas que se hayan instalado hace poco en la región. Otro problema es que dos Floras o Faunas que afecten la misma área (como una Flora o Fauna local y una regional) pueden diferir en sus conceptos taxonómicos o en sus circunscripciones y por lo tanto dar resultados diferentes. Las que están computarizadas poseen muchos de los mismos problemas que las claves en papel.[201] Las actualizaciones se encuentran en las monografías (monographs), un estudio extenso con claves, descripciones y fotos de un taxón en particular, en general en el rango de familia o género, y en las revisiones taxonómicas (taxonomic revisions), similares a las monografías pero menos exhaustivas.[18] Las dos últimas se publican en revistas científicas ([18]:560), y pueden ser accedidas, como las Floras y Faunas, en bibliotecas y hemerotecas con acceso al público de Jardines Botánicos, Museos de Historia Natural y Universidades. Las guías de campo (field guides), que todavía son más fáciles de acceder y poseen muchas fotografías e ilustraciones de las especies de un taxón en una región particular, se han vuelto muy populares y son útiles como una primera aproximación a conocer la flora o la fauna de una región, siempre que se utilicen con cautela y teniendo en cuenta que sólo describen las especies más vistosas y no siempre pueden hacer uso de las herramientas que diferencian un grupo de especies similares.[202][183][168]
Al utilizar una clave, siempre hay que leer cuidadosamente las dos opciones, tener en cuenta todos los caracteres mencionados, no estimar las mediciones sino realizarlas, buscar en el glosario todos los términos que no se conozcan, y tener en cuenta que los seres vivos poseen cierta variabilidad, así que si existe la posibilidad de ver un carácter varias veces (por ejemplo en varias hojas de una planta) se ganará en precisión. ([18]:558) Muchas veces no es evidente que una descripción se corresponde con el espécimen, porque no necesariamente todos los caracteres de una descripción se corresponderán con todos los caracteres del espécimen. Al utilizar una clave es muy probable que en algún momento se llegue a un par de descripciones donde el camino a seguir sea confuso. Cuando es así, hay que seguir los dos caminos, si alguno de los dos es muy incoherente (por ejemplo si pide la observación de varios caracteres que no existen en el espécimen) se descarta. Si no se pueden descartar caminos, se llegará a más de un posible taxón ([168],[18]:558,[20]:496-497). Aunque se obtenga un solo resultado, es importante chequearlo. Una forma de hacerlo es leer la descripción y ver las ilustraciones del taxón o los taxones a los que se llegó, especialmente evaluando la lista de sus caracteres diagnósticos, que es una lista de todos los estados de caracteres del taxón que lo distinguen de los demás taxones. Si coinciden con los caracteres del espécimen, entonces se ha realizado la identificación.([18]:558,[20]:495) Otras formas de hacerlo son la comparación con alguna colección, o en especímenes dificultosos donde aún queden dudas, enviárselo a un especialista en ese taxón particular.([20]:497)
Véase también: Seguir leyendo en Identificación de especímenes, Literatura taxonómica primaria
Historia de la taxonomía
Los orígenes de la taxonomía se remontan a los orígenes del lenguaje, cuando las personas llamaban con los mismos nombres a los organismos más o menos similares, sistema que persiste hoy en día en lo que llamamos los "nombres vulgares" de los organismos. Todas las sociedades humanas poseen un sistema taxonómico que nombra a las especies y las agrupa en categorías de orden superior. El ser humano posee un concepto intuitivo de lo que constituye una especie, producto de agrupar a los organismos según las propiedades emergentes observadas a ese nivel de organización, y esto se ve reflejado en que todas las clasificaciones populares de los organismos poseen remarcables similitudes entre ellas y también con las utilizadas hoy en día por los biólogos profesionales.[203]
La taxonomía biológica como la disciplina científica que conocemos hoy en día nació en Europa. Allí, a finales de la Edad Media se habían formado las primeras universidades, donde se discutían temas de índole filosófica y técnica. Pronto los naturalistas se dieron cuenta de que, si bien los nombres vulgares son útiles para el habla cotidiana, necesitaban un sistema más universal y riguroso para nombrar a los organismos: cada especie debía ser nombrada, cada especie debía poseer un único nombre, y cada especie debía ser descripta de forma inambigua. Al finalizar la Edad Media, con la dominación militar y cultural que una Europa revitalizada sostendría sobre el resto del mundo, el sistema allí consensuado se extendería a todo el globo.
De la taxonomía linneana al origen de los Códigos
Véase también: Taxonomía de Linneo

Portada de la obra de Linneo Species Plantarum (1753),[204] la primera Flora de la taxonomía moderna. Dos años antes había publicado el landmark teórico en el que explicaba los principios con los que construyó esta Flora, Philosophia Botanica (1751).[139] Las dos obras fueron publicadas en latín, el idioma universal de la época.
En 1583, el italiano Andreas Caesalpinus había delineado cuáles debían ser las características de un sistema de clasificación: debía ser fácil de usar y de memorizar, estable en el tiempo, conciso, y además predictivo (ver Greene 1983[205] citado en Judd et al. 2007[18]). Por lo tanto existía la conciencia de que un sistema de clasificación no sólo debía reflejar la naturaleza (ser predictivo), sino también servirle de utilidad a una comunidad de usuarios.


Por un tiempo los naturalistas hicieron intentos de ordenar la información global sobre los organismos y reglamentar sus nombres. Pero sin duda alguna fue en el siglo XVIII que la taxonomía recibió un empujón definitivo gracias al naturalista sueco Carlos Linneo, quien tenía la ambición de normalizar los nombres de todos los animales, plantas y minerales agrupados según sus similitudes morfológicas. Ya había publicado trabajos más modestos antes, pero no fue hasta 1753, ya largamente pasados sus 40 años, que publicó una gigantesca Flora de dos volúmenes en la que se encontraba ordenada toda la información disponible sobre las plantas (incluyendo las sinonimias con los autores más relevantes de los no seguidos), y debido a su éxito, terminó siendo el trabajo que definió el sistema de clasificación que se utiliza hoy en día. El libro se llamaba Species Plantarum ("Las especies de plantas", o "Los tipos de plantas"), y estaba escrito en latín, el idioma universal de la época. En esta Flora Linneo agrupó sus "especies de plantas" en una categoría de rango superior llamada género, llamados por él géneros "naturales", en los que agrupaba las especies que pertenecían a ellos. Al describir una especie dentro de cada género, sólo debía listar los caracteres de la misma que la diferenciaban de las demás especies de su mismo género. Al enfatizar el uso de similitudes morfológicas para agrupar las especies, Linneo estaba clasificando a los organismos en virtud de sus similitudes genéticas, y sin haberlo conceptualizado de esa manero, por lo tanto también evolutivas. En su libro, cada especie era descripta con una frase en latín limitada a 12 palabras, en donde la primera siempre era el género al que pertenecía la especie, y el resto los caracteres que las diferenciaban. A estas pequeñas descripciones, o polinomios, él las llamó "el nombre apropiado de cada especie", pero hizo un importante agregado que había sido inventado en su momento por Caspar Bauhin (1560-1624): la nomenclatura binomial o binomios, como el "nombre corto" de las especies. En los márgenes del libro, seguido del nombre "apropiado" polinomial de cada especie, escribió una sola palabra. Esta palabra, combinada con la primera palabra del polinomio (todavía hoy llamada género), formaba un nombre más fácil de recordar y corto para cada especie. Por ejemplo, la hierba de gato fue nombrada "apropiadamente" con el polinomio: Nepeta floribus interrupte spicatus pedunculatis (en español, "Nepeta con flores en una espiga interrumpida pedunculada"). Linneo escribió la palabra cataria en el margen del nombre apropiado de la especie, que quiere decir "relacionada con los gatos", haciendo referencia a un atributo familiar de la especie. Tanto él como sus discípulos adoptaron el nombre Nepeta cataria para esa planta, y rápidamente sus contemporáneos y los legos que hacían sus propias colecciones empezaron a llamar a las especies del libro con su nombre binomial.
Linneo publicó Philosophia Botanica[139] dos años antes que la Flora global en la que introduciría la nomenclatura binomial de cada especie, Species Plantarum. En Philosophia Botanica compara su nomenclatura con otras en uso en ese momento, e introduce los conceptos taxonómicos que asociará a las categorías de género, especie y variedad. La mayoría de los géneros son heredados de autores anteriores eminentes en cada taxón, convenientemente citados al principio del libro junto con sus obras y las bibliotecas donde podían ser encontrados, y la nomenclatura utilizada para esa categoría es muchas veces el nombre del género tal como lo utilizó la autoridad preferida por Linneo para ese género —si es necesario latinizado o simplificado—, seguido del autor del concepto taxonómico (que no necesariamente era el autor de la primera descripción). Muchos nombres de género fueron utilizados ya en obras de la antigua Grecia y ya se había perdido el autor original, la descripción original o la etimología del nombre. En las últimas décadas cuáles eran los conceptos de "grupos naturales" que debían corresponderse con los rangos de especie, género y familia habían empezado a consensuarse, por ejemplo, en 1694 Joseph Pitton de Tournefort proveyó guías para describir grupos "naturales" de plantas en la categoría de género: los caracteres debían ser reconocibles en todos los miembros del género y ser visibles a simple vista, sin necesidad de un microscopio. En lo posible, debían ser tomados los caracteres de la flor y el fruto, menos lábiles que los relacionados con el hábito. Linneo creía que los grupos que ubicaba en las categorías de especie y género, y algunas familias (grupos de géneros), existían en la naturaleza, eran "grupos naturales", mientras que las categorías más altas sólo fueron utilizadas por conveniencia, formando los grupos según algún carácter visible a modo de clave de identificación. Esta distinción entre un concepto taxonómico "natural" y uno "artificial" no era menor, ya que el creer que los géneros fueran reales (es decir, que existen independientemente de nuestro discernimiento), guía al investigador de una forma diferente de la que lo hace una categoría artificial. En palabras de Linneo:
Los caracteres no hacen al género, el género nos da sus caracteres.
- Carlos Linneo (1751[139]:119)
Con estas palabras, Linneo reconocía que las especies se agrupaban de forma diferente si se elegían caracteres diferentes para diferenciarlas, pero había "algo más" que la mera similitud de caracteres que agrupara a las especies en un mismo género, y había que saber elegir los caracteres a considerar para que la agrupación resultara corresponderse con un "grupo natural". Esta afirmación, fruto de la intuición de Linneo, no estaría asociada a un mecanismo hasta que Darwin pudo publicar su teoría de la evolución en 1859.

Linneo falló en encontrar caracteres que fueran comunes a todos los miembros de una familia, aun en las más "naturales" de ellas, como las umbelíferas.
En la foto: hojas de apio (Apium) con sus pequeñas inflorescencias, hábito de la zanahoria (Daucus), inflorescencias de hinojo (Foeniculum), inflorescencias del cardo corredor (Eryngium), raíz de perejil (Petroselinum).
Linneo agrupó a los géneros naturales en órdenes y clases artificiales, utilizados cual clave de identificación, útiles para que los géneros sean fáciles de encontrar en una lista o un herbario. Los caracteres que utilizó para ello fueron el número de estambres y el número de estigmas de los carpelos, que eran estables dentro de cada género y fáciles de contar. Por ejemplo en la clase Pentandria, orden Pentandria Digyna, ubicó a los géneros cuyas flores albergaban 5 "maridos" y 2 "esposas", quedando agrupadas en el mismo orden las umbelíferas como Daucus (la zanahoria), y la ulmácea Ulmus (el olmo); a las criptógamas, como no tienen flores, las agrupó entre las "plantas de nupcias clandestinas".
Linneo también trató de reconocer algunas familias "naturales", una categoría intermedia entre el género natural y el orden artificial de Linneo (la idea de que algunos grupos naturales podían ser asociados a la categoría de familia data de Pierre Magnol en 1689[206]), si bien falló en encontrar caracteres únicos que pudieran funcionar como diagnósticos de las mismas, y para 1751 ya había reconocido 67 familias (Linneo 1751[139]), dejando muchos géneros sin ubicación. Linneo buscaba caracteres que fueran encontrados en todos los miembros de cada familia, pero su búsqueda falló aún en las más "naturales" de ellas, como las umbelíferas.
El éxito del libro de Linneo se debía no sólo a la claridad de sus descripciones y clasificación sino también a que se había convertido en la revisión más exhaustiva de la literatura taxonómica hasta el momento, con una notación cuidadosa de sinónimos. De esta forma se lo consideraba un "pasaje en limpio" y nuevo punto de partida de la literatura taxonómica, solventando así los problemas de comunicación producidos por la variedad de nombres y sistemas de clasificación locales. A partir de ese momento las actualizaciones de la clasificación hechas por el mismo Linneo o por autores posteriores, intentaron no abandonar la nomenclatura binomial ni el sistema de clasificación impuestos por Species Plantarum.
 Primera página de las especies en Species Plantarum (fragmento). |
En 1758 le llegaría el turno a los animales cuando Linneo publica la décima edición de su Systema Naturae, en el que por primera vez los clasificó como había hecho en 1753 con las plantas, adoptando la nomenclatura binomial. Linneo nombró unas 4.400 especies de animales, incluyendo Homo sapiens, lo que demuestra que la época no tenía reticencias en clasificar al hombre con los animales[nota 15]. Su concepto taxonómico de especie, sus "tipos", eran los que hoy llamamos ecotipos,[139] con diferenciación de nicho (la "esencia" de sus caracteres, o atributos, ya que los de Lineo incluían el factor genotípico y el fenotípico) y localización geográfica, una delimitación diferente de la de la comunidad reproductiva que hibrida en simpatría (lo que posteriormente Mayr llamaría "la especie biológica"), a veces más atomizada y a veces más agregada que la última.
Lineo en Philosophia Botanica (1751[139]) definió el rango de variedad, pero no en la definición con la que hoy lo incluimos en el sistema, no era en palabras modernas un "rango lineano". Metodológicamente, el concepto taxonómico de variedad era el conjunto de plantas tomadas del nicho original de la especie que era criado en condiciones diferentes, obteniendo después de una o pocas generaciones diferentes fenotipos a partir del genotipo original. Las plantas mantenidas en el nicho del ecotipo no pertenecían a ninguna variedad.[139]
| Las variedades son plantas de la misma especie, modificadas por alguna causa adventicia o de otro tipo. El uso de las variedades en jardinería, cocina y medicina hace necesario reconocerlas en la vida ordinaria; de otra manera, las variedades no conciernen a los botánicos, excepto en relación al tema que los preocupa, que las numerosas especies no deberían tener multiplicidad de nombres o ser confundidas. |
Linneo (1751[139]), af. 306, 316. |
En las especies lineanas, el genotipo es invariable, y se encuentra en cada una de sus variedades.
Las excepciones fueron siendo agregadas al sistema de clasificación, autores subsecuentes clasificaron algunas especies en un orden diferente que el resto del género. Desde la aparición de los libros de Linneo a varios autores, como a Michel Adanson (1763-1764[206] citado en Judd et al. 2007[18]), se les hizo evidente que todos los caracteres podían variar y que no hay caracteres "esenciales" para definir a un "grupo natural", que debían ser definidos sólo mediante un conjunto de caracteres tomados de todo el organismo, de los cuales la mayoría, pero no necesariamente todos, estarían en cada organismo de la especie, en cada especie del género, en cada género de la familia. Así se abandonaba el concepto de taxón monotético[cita 64][cita 29] al tiempo que se empezaba a entrar en una era de taxonomía cuantitativa, o politética, con numerosos caracteres combinados en un diagnóstico[cita 65][cita 28][cita 29].
En 1778 Lamarck sugirió que los caracteres utilizados por los científicos para dividir los taxones no necesariamente tenían que ser los mismos que la gente utilizara para identificar algún organismo encontrado. Lamarck promovió separar la clasificación de las claves de identificación, clasificaciones artificiales en las que los usuarios no expertos podían llegar al taxón natural de un organismo dado mediante caracteres que estuvieran a su alcance y con reglas de decisión explícitas. De esta forma se mitigó el conflicto entre los creadores de los sistemas de clasificación, que a medida que aumentaba la información disponible creaban clasificaciones cada vez más crípticas, y los usuarios de esos sistemas, que estaban más interesados en la utilidad del sistema que en la "naturalidad" de sus grupos. En 1789 el exitoso tratamiento taxonómico de Antoine-Laurent de Jussieu, Genera plantarum[209], ubicaría por primera vez por un autor reconocido todos los taxones por arriba de género en conceptos taxonómicos "naturales", iniciando la nomenclatura de las familias de plantas.
El término "Taxonomía" fue propuesto por Augustin Pyrame de Candolle en 1813[210] a partir del griego ταξις (taxis, arreglo o clasificación) y νόμος (nomos, ley), para referir a la ciencia de la clasificación de los organismos.[18] El término "Sistemática" ya se había utilizado p.ej. por Lineo, y proviene del término griego latinizado systema.[211] No sería hasta 1940 en que el término "la Nueva Sistemática" ("the New Systematics", Huxley 1940[212] citado en Mayr et al. 1953) diferenciaría el foco de atención de estos taxónomos de los que continuaban realizando una taxonomía atribuida a la escuela de Lineo de diferenciar "tipos" y debatir la nomenclatura, a los que seguían llamando "taxónomos" o "la Vieja Sistemática" (the Old Systematics).[211]
Un cierto número de sistemáticos, especialmente en Francia y Alemania, adoptó el concepto de "espécimen tipo". Estos "especímenes tipo" eran plantas depositadas en un herbario que representaban la forma más común del taxón, o a veces la forma que el autor consideraba la más "perfecta", de la que el resto del taxón serían plantas degradadas.[18]
La falta de estabilidad nomenclatural en las publicaciones taxonómicas en las décadas siguientes a las obras de Linneo hizo peligrar su logro. Eso llevó al debate y la formación de diferentes Códigos de Nomenclatura, que no eran adoptados por un número significativo de taxónomos. Dos de ellos en particular fueron, en 1813, Théorie Elémentaire de la Botanique ("Teoría Elemental de la Botánica") donde se reglamentaba la elección del nombre válido para cada taxón de plantas; y en 1842, bajo el auspicio de la British Association for the Advancement of Science, el Strickland Code (el "Código de Strickland", Strickland et al. 1842[213]). Éstos serían los que, algo modificados, unas décadas más tarde se consensuarían como Internacionales. (Faltan los cambios en relación a Philosophia Botanica de Lineo.)
De esta forma, desde Lineo hasta los Códigos, nacieron las reglas de nomenclatura zoológica y botánica.
Taxonomía y evolución

Charles Darwin evitó escrupulosamente proponer árboles filogenéticos para organismos reales, para evitar el ridículo que le hubiera quitado mérito a su misión: demostrar la evolución como hecho, mostrando cómo la lucha por la existencia explica la adaptación, y tratando de explicar el cambio evolutivo (transformación) por variación genética y la multiplicación diferencial de genotipos. Al enfatizar cómo la ascendencia común más la divergencia de fenotipos explica los patrones de similitudes y diferencias que nos permiten clasificar organismos en taxones sucesivamente anidados dentro de taxones superiores, Darwin astutamente utilizó un árbol abstracto inmune al ridículo o al error filogenético. Él correctamente arguyó que el plan corporal compartido por miembros de un filo estaba presente en su ancestro común más reciente, y que fue heredado de forma estable generación tras generación, sin cambios fundamentales por cientos de millones de años. Descendiendo en las categorías a través de clases, órdenes, familias, géneros y especies, cada grupo subordinado sucesivo difería de sus relativos más cercanos en caracteres de estabilidad decreciente. ([40]:112)
Darwin en 1859 no fue el primer proponente, pero nunca antes la evolución como hecho había tenido tal penetración en el ambiente científico. Cuando irrumpió a mediados del siglo XIX su teoría de la evolución pronto se admitió, tal como formuló el propio Darwin en El origen de las especies, que el grado de similitud morfológica entre las especies era explicado por la existencia de un árbol filogenético que las conectaba por parentesco de sus ramas ancestrales. Las circunscripciones lineanas asumían que en los conceptos taxonómicos "naturales" de género y especie no existían las variaciones de los caracteres "esenciales". El conflicto entre teoría y colecciones se resolvería cuando se interpretaría que todos los caracteres tienen el potencial de evolucionar dentro de un grupo, y al inicio de la evolución de un grupo nuevo no hay una diferenciación morfológica que justifique la creación de un nuevo taxón.
Un árbol filogenético puede reconstruirse con métodos que involucren técnicas de agrupamiento jerárquicas, no solapadas, como son los grupos que se usan en las clasificaciones "lineanas" (Sokal 1986[2]), por lo que un sistema de clasificación lineano con conceptos taxonómicos "naturales" en principio sería, tal como afirmaba el propio Darwin, congruente con la filogenia de las especies: las relaciones entre taxones en las taxonomías naturales fueron reinterpretadas como el resultado de la evolución, y las categorías taxonómicas como indicadoras de la estabilidad evolutiva de los caracteres que los definen. Sin haber descubierto todavía la estructura del ADN, portador del código genético, que ocurriría recién en los '1960 con Watson y Crick, se acumulaban las pruebas de la herencia de los caracteres probablemente alojados en los cromosomas, para las que los redescubiertos radios de Mendel fueron pioneros, que derivarían en la "Nueva Síntesis" a principios del siglo XX. En pleno auge de la Nueva Síntesis, Ernst Mayr propuso fijar uno de los conceptos de especie que hasta ese momento habían sido llamados la especie biológica —la propuesta por Mayr, para otros autores más neutralmente se llamaría la especie reproductiva— como el grupo de poblaciones capaz de hibridar entre sí y dar descendencia fértil, cuyo pool o acervo genético se mantiene aislado de los de otras en simpatría. E Mayr suele ser citado como un taxónomo que definió el concepto de especie que más fácilmente las delimitaba en vertebrados de tamaño grande —a los que pertenecían los taxones en los que trabajaba—, con una precisión que en esos taxones se corresponde mejor con los demás conceptos que en plantas y microbios.[18] George Gaylord Simpson (1961[15]), descontento con la atemporalidad del concepto de especie biológica de E Mayr, redefinió a la especie como un linaje, una secuencia de ancestros y descendientes con el potencial de fijar un cambio evolutivo en un carácter (lo que Hennig[37] llamaría una apomorfía). Si las especies se definen como poblaciones, o como linajes, no siempre dos organismos de dos especies diferentes se diferenciarán en términos de sus caracteres —por ejemplo, dos poblaciones todavía muy similares en diferentes continentes, cuyo índice de dispersión no les permitió encontrarse por un tiempo prolongado, todavía poseerán al menos potencialmente organismos que no puedan diferenciarse entre sí taxonómicamente más que por la localidad de la que provienen—. Hennig (1950[36] en alemán, 1966[37] en inglés) se encontró con el mismo problema. Asumió que los linajes, que para él también se correspondían con especies, se podían diferenciar taxonómicamente en términos de sus novedades evolutivas adquiridas (apomorfías), por lo que no habría conflicto entre la taxonomía y el árbol de linajes. Linneo (1751[139]) había incluido la localidad de la que provenían los organismos en sus conceptos de especie, y también el fenotipo con el que se encontraban en estado silvestre. La localidad y el fenotipo no volverían a ser incorporados a las clasificaciones.
La incorporación del concepto de evolución a los taxones supraespecíficos fue al principio modelada de forma errática. Por ejemplo, algunas clasificaciones se componían de taxones basados en filogenias que todavía contenían miembros vivientes, como los árboles de Charles Edwin Bessey (1845-1915) y los de Arthur Cronquist (1919-1992). En estos árboles no había ancestros extintos ([18]:43). En la clasificación de Rolf Dahlgren, los ancestros eran sólo miembros extintos, el árbol filogenético era un árbol tridimensional, y los miembros vivientes podían representarse por un corte de un plano de ese árbol, en un diagrama llamado "Dahlgrenograma" con relaciones similares a las de los países en un mapa. Un paso crítico fue la adquisición de los métodos de reconstrucción filogenética atribuidos a la escuela que hoy llamamos cladismo, para la cual el biólogo Willi Hennig fue el que hizo el más valioso aporte en la década de 1960. Hennig modeló que las especies se conectaban entre ellas a través de especies ancestrales –en su modelo original siempre extintas–, y que podía rastrearse la transformación de especies ancestrales a especies derivadas a través de las apomorfías generadas durante una cladogénesis: los cambios de carácter en cada línea filética luego de la divergencia de un linaje en dos. Estos árboles, hoy llamados cladogramas, fueron propuestos por Hennig como material para construir los taxones, que siempre debían ser, a su criterio, monofiléticos. Estos cladogramas tenían el no trivial supuesto de que la historia evolutiva de un grupo siempre ocurre por procesos de cladogénesis (divergencia en dos y adquisición de apomorfías). En la misma década la escuela fenética, para la que el libro de RR Sokal y PHA Sneath (1963[47]) fue fundacional, creaba métodos numéricos de análisis de datos intentando que fueran "ciegos", sin hipótesis de evolución previas (sin hipótesis previas de cuáles serían las apomorfías y las plesiomorfías). Por la época en que Hennig se popularizaba y en respuesta al cladismo la escuela tradicional, que incorporó los métodos de construcción de dendrogramas Hennigianos y los métodos numéricos de Sokal y Sneath en la búsqueda del árbol filogenético, pero difería en cuál es el dendrograma que debe trasladarse a una clasificación, fue llamada evolucionismo, la escuela evolucionista propone, como antiguamente, que puede haber grupos parafiléticos "naturales" que deben formar parte de una clasificación formal. El primero en utilizar el término "cladismo" en reemplazo del confuso término Hennigiano "sistemática filogenética" fue E Mayr (1965[39]), pero ya se utilizaba "clado" en un sentido más amplio, según Hennnig era la hipótesis de ser todos los derivados de una secuencia de ancestros y descendientes con un ancestro común, es decir una hipótesis de grupo monofilético, otros métodos de reconstrucción filogenética también encontrarían "clados" sin determinar cuáles eran las apomorfías y las plesiomorfías. Hennig también redefinió monofilético, que hasta ese momento se asumía incluyendo los grupos monofiléticos y parafiléticos sensu Hennig.[40]
Hacia fines de la década de 1970, cladistas como G Nelson[5], NI Platnick[5] y C Patterson[6] evolucionaron el pensamiento cladista hacia un "cladismo de patrones" o pattern-cladism o transformed cladistics, el cual separaba teóricamente los árboles filogenéticos, es decir los árboles de secuencias de organismos ancestros y descendientes, de los árboles de apomorfías, que pueden haberse correspondido o no linaje por linaje con el árbol de organismos.[cita 2]
Como Hull (1979[3]) ya ha hecho notar, los cladistas en realidad clasifican caracteres en lugar de organismos, tienen que hacer la suposición arbitraria de que nuevas apomorfías se originan cada vez que una línea [filética] se ramifica de su línea hermana. Esto es poco probable en la mayoría de los casos.
E Mayr (1981[4]) citando la nota 28.
Las clasificaciones cladistas no representan el orden de ramificación de grupos hermanos, sino el orden de emergencia de caracteres derivados únicos.
DL Hull (1979[3]) como citado en E Mayr (1981[4]) nota 28.
Nosotros utilizamos el cladograma en el sentido de un sumario jerárquico de los datos de caracteres, no contiene necesariamente supuestos sobre, ni hace predicciones de, evolución y especiación.
Nelson y Platnick (1981[5]:14).
Los sistemáticos siempre han sido, son, serán, y deberían ser tipologistas.
Nelson y Platnick (1981[5]:328)
Formalización de las normas: los Códigos
El que originalmente fuera Théorie Elémentaire de la Botanique en 1813 fue reemplazado en 1930 por el altamente consensuado International Code of Botanical Nomenclature (Código Internacional de Nomenclatura Botánica, ICBN), que trata sobre la nomenclatura de plantas y hongos. El que originalmente fuera el Strickland Code (1842), que reglaba los nombres de los animales, fue revisado y renombrado como International Code of Zoological Nomenclature (Código Internacional de Nomenclatura Zoológica, ICZN) en 1901. El Principio básico de los dos Códigos era el Principio de Prioridad, que promovería su objetivo básico, su principio más[214] básico: la estabilidad en la nomenclatura. El de Botánica estableció el año de la publicación de Species Plantarum, 1753, como el año en que se inicia la nomenclatura botánica moderna, dejando sin efecto los nombres publicados para las plantas antes de esa fecha. El de Zoología estableció el año 1758, el año en que Lineo publicó la décima edición del Systema Naturae y el primero en que utilizó su nomenclatura binomial para los animales, como el de inicio de la nomenclatura zoológica, dejando sin efecto los nombres publicados para animales antes de esa fecha. Los dos Códigos compartieron los mismos "principios de Nomenclatura", y desde su aparición se realizaron periódicamente Congresos Internacionales para su revisión.
Los bacteriólogos, que se habían atenido al Código Botánico, con el tiempo desarrollaron su propio Código (Código Internacional de Nomenclatura de Bacterias, ICNB, publicado en 1958), el único de la taxonomía biológica que posee un registro central desde 1980. El Código Internacional de Plantas Cultivadas a su vez escindió los cultivares de las variedades botánicas en 1952, con su última publicación en 2009, también posee una serie de Registros de Cultivares y sus nombres son invariables, es decir el "tipo" es todo el taxón.[215]
Historia reciente
Desde los '1990 en que empezó a ser "tecnológicamente correcto"[24] realizar análisis moleculares de ADN, la sistemática en sus análisis agrega una cantidad de caracteres de líneas de evidencia diferentes que pueden considerarse bastante independientes entre sí: los caracteres del ADN y ARN, otras moléculas como isozimas, flavonoides..., y caracteres de ultraestructura y micromorfología tomados con los microscopios electrónicos de transmisión y de barrido, además de los avances en paleontología. Estas matrices con una cantidad enorme de datos pueden analizarse con los métodos de análisis filogenético para los que la escuela fenética fue pionera, aprovechando el uso de computadoras, que han producido en las décadas del '90 y 2000 árboles nunca antes tan consensuados y muchos de ellos hasta inesperados, lo cual ha obligado a modificar las clasificaciones al uso, deshaciendo grupos de larga tradición para definir otros nuevos.
Desde los '1990 que los editores publican con preferencia sistemas de clasificación "cladistas" en colaboración de una larga lista de autores: para las angiospermas el grupo que se ha llamado APG (1998, 2003, 2009[216]), para los taxones superiores Adl et al. (2005[87]), etc.
Entre los taxónomos, todavía perdura el debate de si deben aceptarse grupos parafiléticos o no en las clasificaciones. Los grupos parafiléticos y monofiléticos con el mismo ancestro están compartiendo el mismo "tipo" en los Códigos, lo cual lleva hoy en día a una gran inestabilidad en la nomenclatura. El PhyloCode[217], cuyo paper seminal puede rastrearse[217] hasta 1990 (De Queiroz y Gauthier 1990[218]) y cuyo primer borrador fue publicado en 1998, propone agregar las hipótesis cladísticas al "tipo" de los Códigos en uso para restringirlo. El PhyloCode propone nombrar formalmente sólo los grupos cladistas, y que las categorías taxonómicas no se formalicen.
A lo largo del tiempo fueron constantes las propuestas de armonizar, unificar, o reemplazar los Códigos, pero ninguna lograría un alto consenso (Dall 1877,[219] ver también Savory 1962[220] citados en Judd et al. 2007[18]). El BioCode (primer borrador en 1997, segundo borrador en 2011) es la propuesta más reciente de armonizarlos, y desde 1997 se han visto unos pocos cambios en los Códigos en uso en esa dirección.[151] En una versión que se leía sobre el BioCode en el 2015, éste proponía agregar más especímenes de referencia al "tipo" para restringirlo. Otras propuestas están relacionadas con el uso de las nuevas tecnologías informáticas y de redes para publicar nombres y crear un registro central, y sobre la eliminación de las homonimias y la armonización de la nomenclatura de protistas.[151]
Relación con las taxonomías populares
Sabemos muy poco acerca de la relación entre los naturalistas profesionales y los aficionados y el público en general, si bien estas relaciones forman parte del trasfondo histórico que ayudó a modelar la biología sistemática (Judd et al. 2007[18], pero ver por ejemplo Allen 1976[221] allí citado). Sabemos que a medida que aumentaba el número de caracteres con los que se construían las clasificaciones, los taxónomos profesionales en las centurias subsiguientes a la publicación de los libros de Lineo empezaron a tratar despectivamente la taxonomía lineana que todavía era la preferida por el lego, a la que consideraban "una diversión para mujeres y niños".[222]
Como la sistemática tuvo su origen en Europa, los primeros grupos que se formaron fueron aquellos que eran obvios en la flora europea. Además, la disciplina estuvo dominada por europeos durante centurias. Fuera de Europa, especialmente en aquellos lugares que fueran colonias europeas, la dominación europea fue especialmente evidente en que las floras de los países fueron escritas por botánicos europeos y basadas en material depositado en instituciones europeas (Judd et al. 2007[18]:41). Fuera de Europa, los científicos muchas veces desvalorizaron los conocimientos que tenían sobre las especies los nativos que poseían un profundo conocimiento de la Naturaleza en el lugar del que eran originarios, como sucedió con los pueblos originarios americanos.
| «Los nombres nativos parecen perder toda importancia y repetidamente los expedicionarios los señalan como nombres ilegítimos. Para Antonio de Ulloa, "el lenguaje Quechua de los Incas se aproxima más al lenguaje de los niños". Al parecer, los nativos no comprendían las palabras y los conocimientos propios de cualquier sociedad civilizada como Dios, virginidad o inmaculada concepción. Los indígenas americanos tenían innumerables nombres para plantas pero no una única palabra que se pudiera traducir como árbol. Culturas cuya supervivencia dependía del conocimiento y uso de la vegetación circundante, reconocían numerosas plantas de utilidad, sabían cuáles eran sus usos, y les habían dado nombres descriptivos. Sin embargo, es obvio que no compartían con los naturalistas europeos [sus] conceptos de especie, género o clase. Para conquistar plantas extrañas, el europeo se deb[ía] deshacer de contingencias locales y fabricar "tipos ideales" conformes con el sistema de clasificación europeo.» | ||
— Nieto Olarte (2003[223]) | ||
Pocas veces los conocimientos locales han sido utilizados con criterios antropológicos o para la investigación farmacéutica. La comparación de las taxonomías tradicionales con las científicas ha sido objeto de algunas investigaciones.
| «Los indígenas a veces tienen nombres especiales para diversos tipos de Yagé, y les atribuyen distintos efectos, incluso creen que tienen diferentes composiciones químicas. Las clasificaciones hechas por los indígenas pueden referirse a la edad, a las diferentes partes del bejuco, o a formas ecológicas derivadas de variadas condiciones de crecimiento, dependientes del suelo, la humedad, el sombrío...» | ||
— (Muelas 1997, "La Patente del Yagé"[224]) | ||
Perspectivas actuales de la ciencia de la Taxonomía
En las décadas de 1990 y 2000 se ha vuelto a poner de moda la ciencia taxonómica en el ambiente científico,[138][24][225][226] debido en parte al revolucionario enfoque posibilitado por la cantidad de datos derivados del análisis de ADN a los problemas taxonómicos,[138] y en parte a la conciencia de su utilidad, dada la crisis de biodiversidad que estamos viviendo. Nuevas tecnologías relacionadas con la digitalización de los datos y la utilización de internet ofrecen un potencial sin precedentes para la diseminación de los productos de la investigación, pero al mismo tiempo que éstas emergen se observan "vigorosos intentos" hacia el encapsulamiento y el establecimiento de nuevas barreras para la comunicación.[227] Las nuevas herramientas disponibles generan un debate acerca de la utilidad de las reglas de la nomenclatura en su estado actual, y plantean la necesidad de reformar los Códigos, reformas que aún se están debatiendo y algunas de ellas ya se han incorporado.[151]
El impedimento taxonómico

En el período en los '1990 en que fue "tecnológicamente correcto"[24] realizar análisis moleculares de ADN se presupuestó a la Biología Sistemática, pero la descripción de la biodiversidad —de la que el mayor aporte continúa proviniendo del número de puestos de trabajo estables dispersos balanceadamente en todas las regiones en universidades, museos y jardines botánicos—,[23] quedó relegada. ([24][228] citado en[23])
Quizás sorprenda que la descripción de las especies hoy se calcula realizada sólo en un 1% o hasta un 15%, porque las plantas y los animales grandes de regiones bien pobladas como las regiones templadas están relativamente bien descriptos, pero fuera de ellas, como en los estratos arbóreos más altos de las selvas tropicales o en el mundo microbiano con sus tiempos generacionales tan pequeños que lo vuelven vasto y lo hacen evolucionar a una velocidad mucho mayor que las plantas y animales de los que acostumbramos a vernos rodeados, las descripciones tienen actualmente una resolución muy baja. Como muchos autores nos recuerdan, las descripciones de especies "nunca están completas",[229][24][100][230] ya que los descubrimientos de nuevos caracteres y nuevas poblaciones que trae cada nueva línea de evidencia y cada nueva colecta obligan a retestear las hipótesis y chequear cómo evolucionaron los linajes y cómo se dispersaron desde la última colecta. Las demás ciencias de la vida (life sciences) se ven limitadas cuando las hipótesis taxonómicas de las que hacen uso son poco resueltas o confiables,[176][24][231][232] lo que vuelve los objetivos de esta ciencia hoy en día muy alejados de la demanda,[233] a lo que se refiere desde[234] la Convention on Biological Diversity de 1995 como "el impedimento taxonómico".[229][233] Las causas del impedimento fueron llamadas por los científicos "la crisis taxonómica",[229][99] o también "el impedimento a la taxonomía",[233] en referencia a la relativa[233] falta de recursos trasladados a alfa-taxonomía y a los impedimentos sociales[235][236] para su desarrollo, una ciencia que debe competir por los recursos limitados en universidades y subsidios con las mismas ciencias que la tienen de impedimento.[176][230]
Los taxónomos descriptivos tienen pocos gastos además del salario, los que necesitan para publicar, para realizar algún viaje y quizás algún equipamiento adicional, de lo que lo más oneroso es el "molecular lab" del que basta con uno cada 10 laboratorios.[23] A lo que los taxónomos se refieren por recursos en alfa-taxonomía es presupuestar más cantidad de taxónomos con subsidios, plazos y evaluaciones curriculares más realistas, y el mantenimiento a largo plazo de las colecciones:[237]
- (1) Las colecciones[238][230][237] a las que hoy se suman las de archivos digitales y las de cultivos vivos de bacterias, protozoos, algas y hongos,[239] que se sugiere que se depositen en instituciones dedicadas como museos y jardines botánicos. Algunos de ellos poseen colecciones ya centenarias que preservan los ejemplares de la distribución de las especies en otras épocas y son una referencia de su evolución,[236] y son independientes de los arbitrios de un investigador o "pequeño lab"[cita 66] que quizás se deshaga de las colecciones después de la muerte del especialista, o quizás él mismo deba deshacerse periódicamente de las colecciones que no utiliza para hacer lugar en el espacio limitado que le es asignado en la universidad para depositar colectas nuevas. Hoy se estima que se encuentran depositadas entre 1,2 y 2,1[240] o 3 mil millones[241] de unidades. Las colecciones dedicadas deben estar preparadas para el crecimiento constante de sus colecciones[24] de lo que se ocupan en parte sus curadores mediante los intercambios con otras colecciones, quienes también hacen el valioso trabajo de re-identificación de especies para garantizar su actualidad y evitar propagaciones de errores;[238][230] y
- (2) El entrenamiento y salario de nuevos taxónomos quienes, con los datos crudos tomados de sus colectas y otras previamente depositadas en colecciones, publicar trabajos de literatura taxonómica primaria (sólo prepararlos, la finalidad del impresionante programa PEET,[234] no funcionó porque luego no consiguieron trabajo.[237])
Como la mayor parte del trabajo científico, el trabajo del alfa-taxónomo es conducido en proyectos relativamente pequeños, con un investigador principal con un laboratorio asignado en la universidad y dedicación part-time al proyecto, y quizás dos o tres estudiantes de doctorado que ya se han ganado esa primera beca utilizando el laboratorio de otro o investigadores part-time del staff.[242] En taxonomía descriptiva los proyectos son particularmente pequeños, normalmente se hace referencia a un solo investigador, "el taxónomo especializado en tal taxón" en la región dada, que colecta y diferencia especies en solitario con su salario cobrado en la universidad o el museo.[230] La mayor parte de los subsidios (grants) otorgados por agencias gubernamentales o internacionales dedicadas a la manutención de la ciencia son otorgados a la "pequeña ciencia" (small science), con una duración promedio de 3 años,[242] los investigadores mantienen su estabilidad laboral consiguiendo empleo en una universidad —o Museos de Historia Natural, Jardines Botánicos—, a los que dedican la otra parte de su tiempo de trabajo.[242] En ciencia, pocos proyectos son ambiciosos e involucran docenas de científicos, normalmente enfocados en la toma de datos relativamente homogéneos y en cantidad gigantesca organizados alrededor de alguna instrumentación cara y dedicada,[242] la llamada "big science" que aporta "big data", de la que los ejemplos que suelen mencionarse son la astronomía y la física de las partículas de alta energía.[236] Las universidades perciben un porcentaje de los subsidios adquiridos por sus investigadores y del prestigio que éstos acarrean por lo que tienen preferencia por ocupar investigadores que los obtengan.[235] Las agencias de subsidios obligan a la alfa-taxonomía a competir con otros proyectos para ganar los grants, pero ésta termina relegada por razones en apariencia emocionales[24][23] —muchos usuarios y hasta científicos ven a la alfa-Taxonomía o Taxonomía Descriptiva como una "ciencia polvorienta"—,[243][244] y no pareciera que los desperdigados taxones de los que se ocupa su piso pudiera afectar el trabajo del resto de la facultad.[230]
Los investigadores que ya tienen su propio laboratorio son los que eligen de entre los estudiantes de sus materias a los que pedirán un subsidio y un puesto de trabajo y que potencialmente competirán con el mismo investigador que les está dando una oportunidad, esto ha sido mencionado como impedimento,[235][245][230](en rta. a[234]) pero no hay diferencias en ese punto entre las demás ramas de la ciencia y la alfa-taxonomía.([230][235][245]) El mecanismo para evaluar el trabajo de un científico tradicionalmente es su visibilidad,[246][244] de esa forma se promociona la calidad y se desalientan las conductas inadecuadas.[246][247] Pero hace décadas que se lo evalúa en lo que se convirtió uno de los puntos más sobresalientes en su currículum:[230][248] la cantidad de publicaciones en revistas científicas y el prestigio de cada revista en la que se ha publicado medido en forma de "factor de impacto", este último calculado por el Institute of Scientific Information (ISI), desde los '90 parte de Thomson Reuters[nota 16]. El factor de impacto como actualmente calculado es, en cada año determinado, de las revistas indexadas en la Web of Science (WoS) —índice mantenido por Thomson Reuters—, la cantidad de veces que se han citado durante el año los artículos publicados en esa revista en los 2 años anteriores a ese, dividido el número total de artículos publicados por esa revista en esos dos años[237].[250] Los científicos intentan publicar en revistas de alto impacto para alimentar su currículum, pero éstas suelen rechazar los trabajos de índole taxonómica debido a que ocupan mucho espacio en la revista y argumentan[251][252][237][253] que su baja cantidad de citas en los 2 años siguientes les disminuirá el factor de impacto. Las publicaciones taxonómicas son largas, requieren muchas ilustraciones, y típicamente tienen una audiencia muy especializada que dura por muchos años: la vida promedio de una monografía taxonómica puede calcularse en décadas o hasta centurias, en lugar de en meses o en años como el común de las publicaciones.[254][237] Al autor a su vez le toma años o incluso décadas la realización de una monografía;[248] sus trabajos más acabados los publican al final de su vida.[23] Parte fundamental aunque implícita de la estructura de un trabajo científico, artículos muy diversos utilizan por muchos años la información o las claves recogidas de cada publicación de alfa-taxonomía, pero no hay costumbre de citarlos en las referencias (algunas revistas requieren que al nombrar un taxón se cite el autor del nombre y la primera descripción, pero tampoco requieren que se los cite en las referencias).[253][237][244][248] La información recogida impresiona como si no necesitara un autor que la respalde, no hay interés de las editoriales que corren tras las métricas de factor de impacto en citarlos[245], ni una vez publicados se agrega la cita correspondiente ni los lapsos temporales adecuados en los índices de los que se hacen los análisis de rankings, por lo que los autores de las descripciones y claves quedan fuera del rango reconocido de publicaciones en lo emocional y en las métricas de factor de impacto.[248] La necesidad de cumplir con un umbral de impacto, que no mide la calidad ni la necesidad de la descripción de un taxón, sino la "atención"[255][235] que se le presta al mismo, también aumenta la disparidad en las descripciones taxonómicas: ha provocado la deserción en sub-campos de investigación pequeños o impopulares[256], por lo que los llamados "taxones subrepresentados", pobremente descriptos, todavía son ignorados[237][256]; en el otro extremo se le está dando una importancia desmedida, debido a su popularidad, a los estudios que testean las hipótesis taxonómicas exclusivamente por métodos moleculares y a las publicaciones sobre los muy publicitados ADN barcodes, si bien ésta es sólo una de las herramientas utilizadas por los taxónomos para testear los límites de los taxones y para identificarlos[237]. El número de citas se reduce a las que hacen los pocos taxónomos especializados en el mismo taxón en los dos años siguientes a las esporádicas publicaciones, y a su vez si estos colegas publican en las pocas revistas taxonómicas cubiertas por el índice de Thomson Reuters.[254] Los trabajos publicados para el lego o no intencionados para ser citados, como las guías de campo, no son valorados.[23] Las monografías, floras y faunas parecen un trabajo que se hace con un salario que se pagará de todos modos, no se premian con subsidios los trabajos terminados.[23] El taxónomo no percibe la conveniencia de publicar todo el material que tiene,[234][256][23][257] y una fuente de conocimiento valiosa se pierde con el retiro del especialista, volviéndose atrás en las hipótesis taxonómicas sobre su taxón.[234][258] La discriminación de taxones se pasa de forma más segura de investigadores antiguos a jóvenes de boca a boca durante la colecta,[139] y siempre hay que revisar que sea verdadero cada punto del material bibliográfico disponible[139]. La alfa-taxonomía claramente necesita que se amolde a su medida el modo de evaluar sus publicaciones científicas, en esta "era del factor de impacto",[107] I Agnarsson[237], M Kuntner[237], YL Werner[256], R Page[259], MC Ebach[233][260], AG Valdecasas[261][262][233][260], QD Wheeler[233][260], LR Landrum[23], S Castoviejo[261], LF Marcus[261], MR de Carvalho[260], DM Williams[260], SS Nihei[260], MT Rodrigues[260], T Grant[260], LF Silveira[260], H Zaher[260], AC Gill[260], RC Schelly[260], JS Spaks[260], FA Bockmann[260], B Séret[260], H-C Ho[260], L Grande[260], O Rieppel[260], A Dubois[260], A Ohler[260], J Faivovich[260], LCS Assis[260], PZ Goldstein[260], EAB de Almeida[260], G Nelson[260], JL Payne[248], FA Smith[248], M Kowalewski[248], RA Krause Jr[248], AG Boyer[248], CR McClain[248][244], S Finnegan[248], PM Novack-Gottshall[248], L Sheble[248], CHC Lyal, AL Weitzman[202], T Borsch, P Hernández-Ledesma, WG Berendsohn, H Flores-Olvera, H Ochoterena, FO Zuloaga, S von Mering y N Kilian 2015 manuscrito aceptado[263], JV Crisci[264], F Boero[249], H Wägele[265], A Klussmann-Kolb[265], M Kuhlmann[265], G Haszprunar[265], D Lindberg[265], A Koch[265] y JW Wägele[265], KA Seifert[266], PW Crous[266], JC Frisvad,[266] P Pyšek[252], PE Hulme[252], LA Meyerson[252], GF Smith[252], JS Boatwright[252], NR Crouch[252], E Figueiredo[252], LC Foxcroft[252], V Jarošík[252], DM Richardson[252], J Suda[252], JRU Wilson[252], van der Velde[267], F-T Krell[268], J Ellis,[269] HA Benítez,[270] LA McDade[271], DR Maddison,[271] R Guralnick,[271] HA Piwowar,[271] ML Jameson[271], KM Helgen[271], PS Herendeen[271], A Hill[271], ML Vis,[271] GH Pyke,[272] B Páll-Gergely,[273] P Venu[274], M Sanjappa,[274] a pesar de la respuesta del creador del IF, E Garfield[275]). PA Lawrence nos recordaba en el 2008[245] que la forma más segura de evaluar un trabajo científico es leyéndolo —la forma habitual de hacerlo en el pasado—. El problema de este enfoque es que sólo evalúa en retrospectiva, sobre trabajos que tomaron muchos años de realización.[23]
"Dado que muchas especies [biológicas] tienen rangos amplios y complejos y que los especímenes de la mayoría de ellas están desperdigados en las colecciones de los museos de muchos países diferentes, un testeo verdaderamente riguroso de los límites de cada una no es automático ni fácil." ——QD Wheeler (2004[24] ) |
Algunas conductas hacia la nomenclatura formal pueden reducir la calidad de la información taxonómica y repercutir en el prestigio de la rama en general.[237][99] No hay requisitos mínimos para describir una especie, y algunos autores están de acuerdo en nombrar una colecta de la que no pudo determinarse el taxón al que pertenecía luego de la revisión taxonómica siempre que esté correctamente depositada en una colección accesible por otros taxónomos, como si se fijara un epíteto a un "tipo" para después circunscribir las especies cuando pueda presupuestarse,[276] pero hay autores que denuncian que algunas colectas están siendo nombradas formalmente sin que el paper brinde la posibilidad a los demás biólogos de retomar o testear sus hipótesis, o sin haber hecho una revisión taxonómica exhaustiva que lo justificara.[99][237][277][278] Miles de nombres nuevos se publican por año,[150] de los cuales un porcentaje muy alto, que según el autor y el taxón tratados puede ser estimado entre el 25 y el 75%,[279][280] serán nombres sinónimos de nombres previos de especies más ampliamente distribuidas o más variables que lo originalmente descripto. En plantas angiospermas, quizás el grupo más estudiado, se calcula que hay 5 veces más nombres científicos que especies según los conceptos taxonómicos actuales.[281] No parece que se valoren los data papers, las publicaciones que exponen los datos de las colectas sin cambios en las hipótesis taxonómicas —que a pesar del nombre, incluyen muchas hipótesis taxonómicas: los caracteres, las apomorfías, las especies según cada autor...—,[24] en cambio los papers que contienen actos nomenclaturales como el establecimiento de nombres nuevos parecieran asumirse como una señal honesta de la cantidad de trabajo que los autores se habrán tomado.[282] Los nombres no tienen que estar solamente establecidos, tienen que ser nombres "robustos".[230][24]
| Diferencias entre los datos crudos compartidos por la cristalografía y la geobiología[283] | ||
| características de los datos | cristalografía | geobiología |
|---|---|---|
| tipos y value | ‘datos crudos’: un value mayormente rico en información, de larga vida para reusar el formato de texto ‘CIF’: el tipo de datos más comúnmente compartido | ‘plantilla reducida’ tiene valores promedio para observaciones múltiples: lo más comúnmente requerido por otros |
| formato | datos binarios: imagen CIF: estándar de texto amplio en el campo | Plantilla de Excel |
| tamaño | cada imagen o ‘frame’ = 0.25–1 MB set es de aproximadamente 2400 frames (aprox. 1 GB) archivo de texto >500 kB | <1 MB |
| accesibilidad | repositorios de amplio uso en el campo en los que muchas revistas requieren que se depositen los CIFs herramientas OAI-PMHb se están volviendo disponibles para los CIFs | pedidos directos de los datos, dificultuosos y ad hoc, muchas veces sobre la base de publicaciones |
| propiedad intelectual | modelo de servicio: soluciones de estructura de cristales provistos a los químicos; la autoría de los datos es ambigua, requiere negociación antes de que los datos pasen de manos | variable dependiendo de la fuente del presupuesto: primariamente gobierno e industria; la autoría y derechos varían desde completo copyright a muy limitado; común el embargo de largo plazo |
La autoría de la publicación de los datos primarios asociados a las publicaciones primarias, como fotografías digitales de los caracteres, secuencias de ADN, datos de ocurrencia o de colección y de estados de los caracteres de cada espécimen de referencia (R Page,[259] MJ Costello[284][279][285], B Vanhoorne[284], W Appeltans[284], WK Michener[279], M Gahegan[279], ZQ Zhang[279], PE Bourne[279], E Vanden Berghe[285], X Huang[257], BA Hawkins[257], F Lei[257], GL Miller[257], C Favret[257], R Zhang[257], G Qiao[257], VS Chavan[286][241], L Penev[286], D Hobern[286], P Ingwersen[241], Thomson Reuters 2012[287]), evaluaría al taxónomo con la evaluación de la calidad de los mismos y del trabajo que respaldan[284][279][286]. Los datos crudos son el fundamento del trabajo taxonómico, pero hace muchas décadas que se bajaron los costos de la publicación en papel[279] y a pesar de que revistas electrónicas como Zootaxa y Phytotaxa y repositorios externos con la infraestructura y la capacidad de almacenamiento requeridas han reabierto esa posibilidad[279] al 2015, como en biología en general, la situación no se ha revertido y todavía son raramente publicados,[236] como son raramente citados formalmente.[288][289] Cuando se les pregunta, la mayoría de los científicos contesta que guarda datos no publicados de experimentos publicados y también de experimentos "que no dieron" (que no rechazaban una hipótesis nula y no serían aceptados para publicación), que "descansan en el sector inferior de la cajonera del lado izquierdo de su escritorio", y la mayoría de las veces termina en la basura al momento del retiro del especialista, "aún en los retiros más planificados", porque nadie —ni siquiera el científico— sabe exactamente qué hay en esos medios de almacenamiento y formatos que van quedando obsoletos ya que les falta la documentación adecuada para retomarlos cuando los metadatos no anotados ya fueron olvidados.[242] En las encuestas como la publicada en Huang et al. (2012[257]) o la hecha por Thomson Reuters en 2010 (Fig.1[289]) la mayoría de los científicos estaría de acuerdo en publicar sus datos en internet de forma open access si recibiera reconocimiento por ello en alguna forma similar a la publicación en revistas científicas, como coautoría o citas indexadas en bases de datos utilizadas en cálculos de métricas relevantes como las mediciones de factor de impacto[289][257][246]. Sin embargo, y a pesar de que parece evidente que la publicación de los datos aumenta el número de citas de la publicación científica que hace uso de ellos[290], al 2010 la proporción de datos subidos a GBIF, el repositorio más grande de ocurrencias de especímenes creado en el 2001 para satisfacer un objetivo clave de la Convention on Biological Diversity,[279] bajó año a año en relación a la que se detecta o "encuentra",[279] y de ella sólo fragmentos están asociados a publicaciones científicas, siendo la mayor parte datos de colecciones financiadas por organizaciones gubernamentales[279], y más de la mitad de los que lo habían hecho que contestaron la encuesta denegaban la responsabilidad de la calidad de los datos subidos a ese repositorio[279]. Al 2013 está demostrado que una política que hiciera mandataria la publicación de los datos con un aseguro de su preservación y acceso al momento de la publicación del paper altera este paisaje para los autores,[291] y desde el 2011 se han reforzado políticas en esta dirección por parte de varias editoriales, gobiernos y organizaciones (p.ej. Canadá,[292] USA,[293] UK,[294] la National Science Foundation NSF,[295] Evolution[296][297], The American Naturalist[297], Journal of Evolutionary Biology[297], otras adherentes a la política JDAP,[298][297] PLOS journals[299]). El mismo presidente Obama resaltó en su discurso al estilo inicio de sesiones State of the Union 2012 que la "data management" será una de las habilidades críticas de trabajo en este siglo XXI.[300]
Variadas propuestas se han hecho del tipo de modernizar la infraestructura para mejorar la eficiencia del trabajo y facilitar la evaluación del mismo, algunas de las cuales se verán en las secciones siguientes. La que sobrevuela todas ellas es la de digitalizar y proveer acceso online a los recursos taxonómicos.[280] Se propone digitalizar la literatura en papel,[301][302][303] que las publicaciones sean de contenido libre (ej.[229][304][305], pero ver[306]), que los museos digitalicen sus colecciones[307][305][240],[308] que éstas sean de contenido libre[305][309], que hubiera "megarrevistas" electrónicas para las publicaciones taxonómicas —que pueden llevar más de mil páginas—,[310][311], que éstas sean open-access[311], que se alojen los datos primarios utilizados en las publicaciones en bases de datos públicas para verificación[248], que esos datos estén relacionados con claves interactivas[225][280], que éstas sean accesibles[225], que sean actualizables.[280] Son muchas las propuestas de un único portal que tenga acceso a todas las bases de datos, tanto de colecciones como de bibliografía (p.ej. EOL[281]), a veces asociada a la propuesta de relacionar esos datos en una única clasificación, también se propone relacionarlos mediante buscadores (search engines) especializados que detecten e indexen sus identificadores estandarizados,[312] como son comunes las propuestas de crear listas de taxones establecidos.[313] Tecnologías que se promueven son la digitalización 3D de imágenes de preparados al microscopio (ej[225],rSEM[314]), la creación de fotografías focus stacking de especímenes tipo, ensamblarlas en imágenes 3D[307][314], que los museos compren la tecnología para el análisis de ADN y quizás encontrar barcodes (ej. Tautz et al. 2003[258]), o que haya un "molecular lab" cada 10 laboratorios[23], que se implemente la toma de datos telemétricos y monitoreo satelital de distribución y ecología (Patterson et al. 2006[281]). Estas iniciativas son tecnológicamente sencillas, en este momento las principales dificultades están relacionadas con su integración: la falta de estandarización de los formatos[236] —para lo cual está trabajando el TDWG—, las dificultades en la creación y mantenimiento de registros[151][236], índices, e Identificadores Globales reusables[315] (algunos proponen en bases de datos copiables[315] y cosechables por buscadores específicos[315]), y que la financiación no debe perder el objetivo de obtener descripciones de especies publicadas, que incluyan claves de identificación bien documentadas[202] y accesibles[225][202] que eviten la duplicación de esfuerzos en el futuro, lo que han llamado a veces peyorativamente "la actitud hacia el usuario final" o del usuario final (end-user attitude[316][cita 67]). Se han financiado proyectos "horizontales" en miras de organizar una "big science" a partir de una "small science" articulada,[242] como GBIF (repositorio de ocurrencias y fichas de colección de especímenes) y los proyectos asociados de digitalización de los catálogos de museos y herbarios; y el registro de nombres y actos nomenclaturales Zoobank y algún proyecto de digitalización de catálogos de bibliotecas y hemerotecas; y desde el punto de vista del trabajo del taxónomo, al '2010 ha demostrado en vistas del número real de especies[317] —mucho más alto que el estimado en centurias previas—,[317] organizarse por taxones de tamaño más pequeño que los Reinos gobernados por los Códigos.[151][229] Las monografías son "consumidoras de tiempo más que consumidoras de dólares" además del salario,[23] y los subsidios son pocos y muy grandes y de corto plazo para el equipo y el tiempo que necesita cada taxónomo,[23] por lo que la falta de taxónomos en cada taxón podría solventarse otorgando subsidios de tamaño más realista y con evaluaciones curriculares más realistas[23] (PEET podía haber otorgado 10 veces más subsidios 10 veces más pequeños, a 400 labs en lugar de 40, por los 5 años que duraba el subsidio[23]; la Flora of North America podía haber pagado pequeños subsidios a quienes colaboraron en ella a medida que iban terminando su trabajo[23]), en lugar de ser un equivalente a "ganarse una lotería".[23] En cada taxón de tamaño más pequeño cuando cada cuerpo mandatario internacional lo decida puede organizarse una nueva fecha de inicio de la nomenclatura, la que hoy en día sólo existe en bacterias, fijada en 1980.[151][229]
El copyright ("derecho de copia") es de concernimiento en la preservación de la literatura taxonómica. La literatura taxonómica primaria en una distribución sin formato no se vería afectada por las leyes que restringen la realización de copias y su distribución (p.ej. el Convenio de Berna), pero son pocos los investigadores que publican comentarios al respecto (p.ej. Costello 2009) y parece que las editoriales no se hacen eco del tema.
Únicamente las publicaciones de De Carvalho y colaboradores brasileños denuncian que los "piratas de la biodiversidad" pueden boicotear el trabajo del taxónomo.
El rol del ADN y los "marcadores de ADN"
Los análisis moleculares de ADN se utilizan hoy habitualmente en los análisis filogenéticos y desde los '90 que unos pocos cientos de bases, con su cantidad enorme de combinaciones potenciales, parecen bastar para hacer análisis de identificación y parentesco.[138] Esto llevó a algunos autores, como Hebert et al. (2003)[318] y Tautz et al. (2003),[258] a proponer asignar un rol central al ADN en la descripción e identificación de las especies, de forma que una muestra de ADN y la lectura de una de sus secuencias de bases debería ser una especie de marcador para ubicar el taxón al cual pertenece un espécimen, de una forma similar a como se usa el código de barras en los supermercados. Tautz et al.[258] incluso proponen reformar los Códigos para que este "identificador" sea un carácter agregado al "tipo". Esta "Taxonomía basada en ADN" aún adolecería de muchos de los mismos problemas que tienen las demás propuestas de identificaciones o de agregar atributos al "tipo", en particular, hay que decidir qué secuencia usar, ya que algunas secuencias no dan información que diferencie al taxón de los demás. Esto puede ser porque un mismo gen puede mantenerse inalterado durante millones de generaciones y muchas especiaciones después de su formación, o debido al fenómeno de introgresión, de forma que un gen que se había diferenciado vuelve a su estado anterior. Por lo tanto, de la misma forma en que no es conveniente confiar en un solo carácter morfológico para identificar una especie, tampoco es conveniente confiar en una sola secuencia de ADN. Además, aun cuando la "Taxonomía basada en ADN" fuera financiada, habría que preguntarse si es necesario agregar un requerimiento extra al ya lento proceso de describir nuevos taxones, en especial teniendo en cuenta que se calcula que sólo el 10 % de las especies del planeta ha sido descrito. Debido a eso, probablemente la mayoría de los biólogos verá las secuencias de ADN como un complemento de la información morfológica, más que intentar reemplazarla buscando verdaderos barcodes. De todas formas, los Códigos no especifican ningún carácter en particular para diagnosticar nuevos taxones, así que la "Taxonomía de ADN", con el tipo nomenclatural de costumbre, ya es válida, aunque la identificación mediante caracteres visibles puede ser de uso más inmediato y su realización más económica que la lectura de un verdadero barcode.[138][202][319][320][321][264]
| Los principales[322] recursos de "barcodeo de ADN" (DNA barcoding) al 2012[322]. | ||
|---|---|---|
| . | ||
CBOL (Consortium for the Barcode of Life) (enlace roto disponible en Internet Archive; véase el historial y la última versión).. | 2004 en adlnte. | Presidente: Dr. Scott E. Miller, opera desde el museo Smithsonian Institution’s National Museum of Natural History en Washington. |
| Realiza barcodes para 200 organizaciones en 50 países. | ||
| . | ||
BOLD (Barcoding of Life Data System). | 2005 en adlnte. | Opera desde el Biodiversity Institute de Ontario, Canadá, cuyo director es el Dr. Paul Hebert. |
| Opera como banco de trabajo online: repositorio de barcodes, análisis, interfaz para submisión a GenBank, herramienta de identificación de especies, conectividad con recursos externos. | ||
| . | ||
iBOL (International Barcode of Life). | 2010 en adlnte. | El hub científico es el Biodiversity Institute de Ontario y el director científico es el Dr. Paul Hebert. |
| Compromisos y grupos de trabajo sin fines de lucro en 25 países "tanto desarrollados como en desarrollo". | ||
| Los atributos estandarizados por el "barcodeo de ADN" apoyado por el CBOL.[323] | ||
|---|---|---|
| . | ||
| Eucariotas, principalmente animales | Una región de 658 pares de bases del gen mitocondrial cytochrome c oxidase I (COI). | Hebert et al. (2003[318]) |
| . | ||
| Plantas | Dos fragmentos plastídicos de 500- a 800- pares de bases de la subunidad grande del gen ribulose 1,5-biphosphate carboxylase (rbcL) y del gen maturase K (matK) | Hollingsworth et al. (2009[324]) (CBOL Plant Working Group 2009[324]) |
| . | ||
| Los atributos a estandarizar por el "metabarcodeo de ADN".[323] | ||
ADN ambiental (no separado por organismo) | Secuencias a estandarizar luego de utilizar la tecnología NGS (next-generation sequencing) para referenciar los genomas organelares completos y el rDNA nuclear repetitivo de todas las especies. | Taberlet et al. (2012[323]) y referencias en ese texto |
Al 2003 ya era rutina que los microorganismos se delimitaran con métodos moleculares,[138] y está claro que para dilucidar el árbol de la vida completo, sería útil secuenciar los mismos genes en muchos taxones diferentes.[138] Para lograr esto, sería necesario un "proyecto genoma horizontal"[138] y un sistema de archivo de secuencias de ADN como el INSDC[325] que incluye al estadounidense GenBank, más allá de si el ADN se vuelve un requerimiento en la descripción o identificación de especies o si se lo agrega al "tipo".[138] Al 2005, dos proyectos habían recibido el millonario presupuesto para llevar a cabo la empresa global de secuenciación de barcodes[320][100] —nombre que quedó a las secuencias seleccionadas por los proyectos presupuestados—, el Barcode of Life Data System[326] (BOLD)[327], y el Consortium for the Barcode of Life[328] (CBOL, "Consorcio para el código de barras de la vida").[322] Sus seguidores promocionan los casos de macroorganismos en que el agregado de esa línea de evidencia molecular a las hipótesis taxonómicas llevó al esclarecimiento de problemas de sinonimias, como cuando la secuencia de ADN "barcode" COI logró diferenciar en 4 especies "crípticas" la sanguijuela medicinal europea.[329][330] Como en ese mismo ejemplo, la secuencia estandarizada de ADN que es parte del gen COI tomada de un espécimen es de utilidad para delimitar especies sólo acompañada de la matriz de caracteres del espécimen tomados de otras líneas de evidencia, moleculares y no moleculares, y de una preidentificación morfológica —QD Wheeler nos recuerda que muchos especímenes de referencia descansan en las colecciones sin una caracterización morfológica adecuada,[24] y en un porcentaje significativo está mal la determinación o no se ha intentado su identificación—,[24][176] sólo de esta forma puede utilizarse esta secuencia en una matriz de análisis filogenéticos;[230] y posteriormente quizás forme parte de una clave de identificación.[99][100][331][137] A diferencia de lo que ocurrió en los '90 en que los "barcodes" se tomaban sólo sobre especímenes morfológicamente caracterizados hasta hipótesis de especies,[18] al 2013 los barcodes se han producido a una velocidad mayor que la revisión taxonómica que debería acompañarlos, lo que ha hecho proliferar en el estadounidense GenBank los llamados taxones "en la oscuridad" (dark taxa).[35] Éstos podrían formar parte de taxones conocidos en los que el organismo del que se extrajo el ADN aún no haya sido ubicado, o de taxones nuevos, no caracterizados.[35] Muchos barcodes "sin nombre" (unnamed) habían sido suprimidos por GenBank, pero aun así su número siguió aumentando en los últimos años.[35]
En respuesta a la intensa propaganda[230] realizada a favor de esta empresa —que para muchos autores desbalanceaba las necesidades presupuestarias de la alfa-taxonomía—,[100][99][24] el término "taxonomía integrativa" fue acuñado, independientemente y casi al mismo tiempo,[122] por Dayrat (2005)[99] y Will et al. (2005),[100] el último luego de una conferencia, el DNA barcoding debate Will vs. Hebert,[100] el "padre del barcode".[243] La taxonomía o aproximación integrativa alude a la práctica tradicional, y la esperable en taxonomía, de integrar líneas de evidencia durante la generación de hipótesis taxonómicas, y en particular a la integración de los variados métodos moleculares con los morfológicos como la única primera aproximación válida para delimitar taxones en la categoría especie.[99][100] Desde esa fecha un considerable[226][122] número de estudios taxonómicos se hizo eco del término.
La presupuestación de los proyectos promovidos por Hebert et al. (2003[318]) dejó como legado lo que algunos autores[323] llaman la primera región de ADN "horizontal"[138] estandarizada para su utilización como atributo en una matriz de análisis filogenéticos, para lo que anteriormente se habían utilizado muchas regiones de ADN nucleares y organelares diferentes enfocadas en taxones de tamaño más pequeño[323] (ver p.ej. en plantas). QD Wheeler decía en el 2004:[24] "Estamos en el umbral del salto más excitante de la 'Taxonomía basada en morfología' [identificación de especies mediante claves morfológicas] desde Linneo (1758), lo que vuelve estos tiempos un momento inoportuno para darse por vencido". ... "Los datos visuales morfológicos nunca habían podido ser eficientemente comunicados en el pasado. Ahora tenemos la tecnología para una edad dorada de la morfología comparativa si podemos sobrellevar las presiones para conformarnos a lo 'tecnológicamente correcto'".
La crisis de biodiversidad
La crisis de biodiversidad causada por actividades humanas, como las que llevan a la pérdida del hábitat y la introducción de especies exóticas, es tal vez comparable en magnitud a las grandes extinciones registradas en los períodos geológicos (PH Raven 2002[332]). El término "biodiversidad" fue acuñado[333] por EO Wilson como un reemplazo eufónico de "diversidad biológica" en una serie de populares ensayos donde desde mediados de los '1980 ponía el foco en ésta y su crisis; en 1992, el tratado internacional llamado Convenio sobre la Diversidad Biológica (Convention on Biological Diversity), ratificado por todos los países miembros de la ONU salvo Estados Unidos, reconoció como un derecho internacional la valoración de la biodiversidad en el contexto de su conservación y uso sustentable. Sin embargo, existe una conciencia general de que no será posible conservar toda la biodiversidad.[334] Los especialistas deben calcular en qué hábitats y en qué especies focalizar los magros presupuestos asignados a la conservación, priorizar los hotspots (regiones con una concentración de especies endémicas con peligro de perder el hábitat[335][277]), y generar modelos predictivos acerca del futuro de la biodiversidad.[336] A 1995, se había hecho evidente para el CBD que los especialistas no pueden medir una biodiversidad no descripta, lo que bautizaron como "el impedimento taxonómico",[229][233] a raíz del cual el organismo impulsó el desarrollo de la alfa-taxonomía. Las medidas concretas encaminadas a cumplir estos objetivos empezaron a ser visibles en algunos países e instituciones internacionales a fines de 1990[309], momento a partir del cual se hizo evidente un segundo impedimento: la elección del concepto taxonómico a utilizar tiene sus consecuencias en las decisiones de conservación. Por ejemplo, Peterson y Navarro-Sigüenza (1999[337]) analizaron la cantidad de especies de aves endémicas de México utilizando dos clasificaciones diferentes basadas en distintos conceptos de especie. Según el concepto biológico de especie o BSC (que agrupa dentro de la misma especie a los organismos con capacidad de hibridar y dar descendencia fértil), obtuvieron 101 especies, con la mayor concentración de especies endémicas en las regiones montañosas del sur y del oeste. En cambio según el concepto filogenético de especie o PSC (que asigna como miembros de una especie a los que poseen evidencia de conservar caracteres en un análisis de filogenia, más allá de si pueden hibridar con otras poblaciones o no), obtuvieron 249 especies endémicas, con la mayor concentración en zonas tanto llanas como montañosas del oeste. La elección de una clasificación u otra, por lo tanto, afecta enormemente las prioridades de conservación de cada hábitat. La estabilidad que presentaron por al menos dos décadas desde las influentes publicaciones de E Mayr donde proponía su concepto de especie biológica hasta ~1985 algunos taxones bien conocidos, contrasta fuertemente con la reciente incertidumbre sobre el concepto de especie a utilizar y la "inflación" en su número, que especialmente en los grupos bien estudiados, incluso en primates, ocurrió en parte significativa por un cambio en la preferencia de los taxónomos del concepto de especie biológica o BSC al concepto de especie filogenética o PSC.[117]
La preferencia de conservación de la categoría especie trajo un problema extra a la clasificación.[117] Esto ha animado la elevación al rango de especie a las poblaciones que necesitaban protección, más allá de si la decisión podía ser respaldada científicamente.[117] El fenómeno fue descrito en la tortuga negra Chelonia agassizi, pero es probable que haya ocurrido también en otros taxones.[117] Esta inflación de especies estaría sesgada hacia formas carismáticas, de gran tamaño, raras y en peligro de extinción, que atraigan interés del público y científico para su conservación.[117] Un interés local puede llevar a una población a ser listada aparte, pero su sobrevivencia puede ser más probable si se la gestiona como un componente de una metapoblación viable (Isaac et al. 2004[117]).
Ha habido otras propuestas de medidas de biodiversidad, con sus consecuencias en las propuestas de conservación, se ha propuesto como medida de biodiversidad la categoría de género (incluso en primates, como en Harcourt et al. 2002[338]) o las poblaciones, otros estudios abandonaron la taxonomía biológica para describir la biodiversidad con medidas de "diversidad filogenética".[117] Sin embargo, esas medidas pueden quitar atractivo presupuestario debido a que la opinión pública favorece los taxones especie como unidad de medida.[117] Una solución sería que los checklists o inventarios (listas de especies por región) se agreguen de forma globalizada especificando el concepto de especie utilizado y se enlazaran a sinonimias bajo otros conceptos de especie,[117] de forma que se sincronicen todas las clasificaciones que afectan una región para presentar su diversidad taxonómica sin dejar de lado ninguna autoridad. La tecnología para hacerlo está disponible, pero se necesita una inversión institucional fuerte y de largo plazo.[117]
Informática de la Biodiversidad

Las zonas estandarizadas en Norteamérica como "regiones TDWG" (2ªed) del Biodiversity Information Standards (anteriormente TDWG, "Taxonomic Data Working Group"). "Si bien muchas veces no lo suficientemente reconocidos, los estándares TDWG son utilizados en muchos softwares populares de biodiversidad informática, cuyo rango abarca desde pequeñas soluciones de escritorio hasta redes internacionales de datos de gran escala."[239]
El campo de acción de la informática de la biodiversidad (biodiversity informatics) son las ciencias biológicas que utilizan como unidad de trabajo los organismos, lo que la diferencia de la ecoinformática (ecoinformatics, la informática al nivel de organización de los ecosistemas) y la bioinformática (bioinformatics, al nivel de organización molecular y fisiológica).[239]
El material tomado como digital o a digitalizar en taxonomía consta de los "datos crudos" y sus metadatos, y de la "literatura taxonómica primaria" y sus respectivos metadatos. La digitalización requiere un id "único" para cada uno de ellos, idealmente "global", es decir único en internet. Por ejemplo la forma más directa de crear un id global es concatenar el id del dato con el de su tabla en la base de datos con el identificador de esa base de datos en internet.[339] Hay varias propuestas de formación de id globales y alguna de ellas puede volverse estandarizada en el futuro, cuando la aceptación por parte de la comunidad de usuarios lo decida. En internet además se busca que este GUI (identificador global único, global unique identifier) pueda "resolverse", enlazarse a través de internet a un archivo con sus metadatos y quizás una lista de enlaces a copias del dato taxonómico, los dos alojados en su base de datos online, en lo que cuando son adecuadamente citados se llamó el mundo emergente de los Linked Open Data de la Big New Biology, la "Nueva Biología Gigante" más "data-céntrica".[236] Las discusiones en la comunidad de la informática de la biodiversidad se enfocan en 3 tipos de GUIs "autorresolubles", es decir que sean una dirección de internet que al tipearla en la barra de direcciones devuelva la página con los datos o los metadatos: los HTTP URIs (es decir URLs), los GUI recomendados como estándar por el TDWG que son los LSID (que hacen uso de los DNS de internet, y resuelven a una página de metadatos), y los DOI, que hacen uso de una autoridad central para resolverse y son por ejemplo los asignados por CrossRef para publicaciones científicas que los resuelve al dato[160]. Los GUIs no ocupan espacio en las publicaciones taxonómicas porque los metadatos y el material al que hacen referencia pueden agregarse en bases de datos externas, las bases de datos deberían ser "copiables" para amortiguar la posibilidad de que metadatos y datos se pierdan, y para independizarse de las autoridades que mantienen las bases de datos mediante buscadores —search engines— específicos o inespecíficos que indexen los GUI, sus metadatos, sus locaciones, sus GUI sinónimos y usos posteriores de los mismos.[35] Como la unidad de trabajo en la clasificación es el organismo, los datos crudos y la literatura primaria deberían enlazarse a los id de los organismos o cepas referenciados en ellos, por lo que implican la necesidad de un GUI por organismo/cepa.[34] Los id de los organismos son el taxón terminal de las clasificaciones según cada autor. Cada nombre de taxón sensu cada fuente también necesita un id, para poder separar sinónimos nomenclaturales del nombre y sincronizar conceptos sinónimos con diferente nombre.[34][35]
Los datos primarios y su literatura asociada pueden agregarse en proyectos "verticales" de un taxón algo superior o con líneas de evidencia que requieren diferentes especialistas, la digitalización de los metadatos primarios que tengan "actos nomenclaturales" (establecimientos de nombres, etc.) por taxón "vertical" en miras de crear las tan esperadas Listas de Nombres Adoptados (no suprimidos) necesarias para los Códigos en uso es considerada una prioridad para el BioCode.[151] También hay proyectos "horizontales" como los de ADN barcodes, la Biodiversity Heritage Library que digitaliza literatura taxonómica, los proyectos que digitalizan los especímenes y sus metadatos (ficha de colección, etc.) de las colecciones de los museos. Algunos sitios web, como la Encyclopedia of Life (EoL) son "iniciativas globales", funcionan como portales que presentan la información "agregada" de "asociados" (partners), las bases de datos indexadas en ellos. El candidato obvio para ser el id que combinara bases de datos diversas había sido el nombre científico, como fue utilizado en iSpecies y EOL, pero pronto se vieron sus limitaciones como identificadores debido a que no son ni completamente estables ni globalmente únicos: en los nombres científicos son permitidas, o inevitables, las sinonimias y las homonimias.[160] Idealmente en el futuro los nombres científicos sean reemplazados por id de conceptos taxonómicos[34][35] (de nombres sensu cada autor), que puedan resolverse hacia la cita a la que corresponde la descripción del taxón utilizada con todas sus referencias.[35]
Entre las iniciativas más populares desde el 2003 quizás estén:[138]
GBIF[340] Financiado por agencias gubernamentales en acuerdos internacionales luego del CBD. Portal de acceso a los datos de ocurrencias provenientes de cientos de instituciones de todo el mundo. Recomienda enviar las colaboraciones individuales a iNaturalist,[341] eBird[342] o la Encyclopedia of Life (EOL).[343]
Species 2000.[344] Al 2015 se lee: "ITIS y Species 2000 se han asociado para formar el Catálogo de la Vida (Catalogue of Life, CoL) que orgullosamente provee el esqueleto de la Enciclopedia de la Vida (Encyclopedia of Life, EOL[345])".[346] La Enciclopedia de la Vida nació en el año 2003[347] y puede agregarse información luego de registrarse como usuario. Es muy publicitada en Science (por ejemplo en Stuart et al. 2010[348]).
Tree of Life,[349] desarrollado en los '90 y anunciado formalmente en 1996,[350] el esqueleto taxonómico lo provee la comunidad científica, la comunidad de usuarios puede agregar contenido multimedia por taxón. Al 2007 se lee, "muchos usuarios y contribuyentes de Tree of Life han notado el extenso solapamiento en los objetivos de EOL y ToL, y ha habido algún concernimiento acerca de la supervivencia a largo plazo de ToL corriendo en paralelo de un proyecto similar pero mucho más grande, financiado a una escala de órdenes de magnitud por encima de los recursos de ToL. En orden de evitar la duplicación de esfuerzos, ToL y EOL han acordado coordinar cercanamente las actividades futuras, compartir contenido y herramientas de software".[351] El proyecto se dejó de actualizar en el 2011.[352]
Pero hubo más de 50 proyectos en todo el mundo.[138] Un problema común a muchos de ellos fueron las expectativas irrealistas de los usuarios respecto al número de datos que debían ser curados, por lo que luego de la fundación inicial los recursos se desvanecían y los proyectos quedaban detenidos.[353] Un ejemplo fue All Species Foundation, creado en el 2001 por un ambicioso grupo de "Silicon-Valley types" entre los que estaban prominentes investigadores como EO Wilson de Harvard, y Peter Raven del Missouri Botanical Garden.[354] Su objetivo era catalogar todas las especies del planeta en 25 años, pero en el 2005 el proyecto había muerto.[354] Se habían terminado las donaciones luego de finalizada la "explosión de la burbuja punto com" (dot.com bubble burst), una visita del 2005 a su página web sólo decía, "Nos fuimos" ("Gone").[354] Ni los assets fueron reutilizados.[354] EO Wilson ya había retomado la idea[355] en su nuevo proyecto, la Encyclopedia of Life (EOL), con el objetivo a plazos más modestos de una página web para cada especie del planeta, presentada por el mismo EO Wilson en el 2003.[356]
Pero si bien ya en el 2010 Padial et al.[226] calcularon que tomaría 400 años de trabajo taxonómico constante llegar a describir las cerca de 10 millones[357] de especies de eucariotas, al 2012, la realidad es que las colecciones de historia natural están casi tan lejos de la digitalización completa como 20 años atrás, y aún no hay una verdadera database central de acceso a colecciones, ni un registro completo de las mismas para ofrecer a los usuarios.[240] A través de GBIF, la fuente más grande de información, Ariño (2010[358]) estimó que eran accesibles no más del 3% de los datos de especímenes preservados en las colecciones de todo el mundo.[240] Blagoderov et al. (2012[240]) calcularon que el costo de la digitalización de los cerca de 30 millones de especímenes de insectos alojados en la colección de entomología del Museo de Historia Natural de Londres, por ejemplo, tomaría 23 años de trabajo continuo de las 65 personas que componen el staff del departamento, y en el mundo, dependiendo de la colección los costos varían entre 0,50 a unos cuantos dólares por espécimen para capturar sus datos completamente, por lo que la digitalización completa de todas las colecciones de historia natural del planeta costaría unos 150 mil millones de euros y tomaría unos 1500 años.[240] La solución más común propuesta es la prioridad de la digitalización sobre la base de la demanda, también se propone financiar los costos de una digitalización "industrial" de las colecciones.[240]
Zootaxa fue la primera "megarrevista"[310] electrónica con referato. Creada en el 2001,[310] llegó a publicar monografías de más de mil páginas[310], y en el 2004 ya se perfilaba como la mayor revista en taxonomía zoológica,[310] momento en el que entró a las mediciones de factor de impacto de ISI.[310][359] Al 2008 se promocionó como la primera en publicar un "paper cibertaxonómico" con enlaces y otras utilidades.[310] A ella siguió una generación de nuevas "revistas rápidas" en taxonomía,[310] incluyendo Phytotaxa (2009[360][361]) y Zookeys (2008[314]). Pensoft (Zookeys, Phytokeys, Mycokeys) distribuye desde el 2011 de rutina los datos taxonómicos publicados hacia una serie de recursos online que incluyen GBIF, EOL, y la indexación en Zoobank,[362] al 2015, una actualización del software permite que estas bases de datos externas "cosechen" los datos de los papers que corrigen datos de papers anteriores, así los datos se corrigen dinámicamente en ellas.[362] Estas revistas también hicieron las primeras pruebas de publicar papers "de datos" (data papers).[286] En el 2012 los dos Códigos permitieron la publicación electrónica de actos nomenclaturales, facilitando el uso de este recurso.[310][363][360]
Algunas revistas tradicionales hoy exigen que los datos crudos (raw data) sobre los que fueron tomados los datos de publicaciones taxonómicas sean alojados en bases de datos públicas.[248] También son usuales los artículos "suplementarios" publicados en formato exclusivamente electrónico.
Por otro lado, al 2014, las bases de datos externas ofrecen la posibilidad de publicar monografías taxonómicas fuera del circuito de las revistas científicas, con las facilidades que podría dar una base de datos interactiva.[280][263] Algunos softwares para monografías son las plataformas Scratchpads, KE-EMU, Diversity Workbench Platform, EDIT Platform for Cybertaxonomy (el último el único de los listados con capacidad de alojar clasificaciones alternativas), softwares para alojar una base de datos descriptivos y crear claves son Lucid2, Xper2.[263]
La armonización de los Códigos y el BioCode
- Artículo principal: BioCode
La IUBS es un organismo que tiene su participación en los Códigos en uso y en 1985 formó un Comité de Nomenclatura Biológica, el cual desde ese momento ha organizado numerosos congresos y reuniones, a los que se han sumado el auspicio de la IUMS (International Union of Microbiological Societies) y del IAPT, y el apoyo de organismos como CAB International, el Linnean Society of London, la Royal Society of London, entre otras. En 1994 se formó el IUBS/IUMS International Committee on Bionomenclature (ICB). Fue el ICB quien propuso un primer borrador de un Código unificado de nomenclatura después de sus esperables congresos y revisiones que fue presentado como el "Borrador del BioCódigo 1997" (Draft BioCode 1997) (DL Hawksworth, presidente de la IUBS).[151]
El Biocode propuso que fuera utilizado en reemplazo de los Códigos en uso, pero su uso no fue implementado por los autores de publicaciones taxonómicas. La crítica principal consistía en que había baja disposición de botánicos y zoólogos en aprenderse todo un conjunto de reglas nuevas,[364] argumentando que los problemas existentes podían superarse reformando cada Código en uso, lo que quizás está relacionado con la crítica de haber sido "poco maduros"[365][366][367][368] al presentar una propuesta semejante.
La crítica fue escuchada y la nueva versión del BioCode, el Draft BioCode 2011, ha sido sustancialmente reescrita de forma no de reemplazar los Códigos actuales, sino para proveer un marco común que los englobe. Está repensado para ser utilizado junto con los Códigos en uso —a los que llama los Códigos Especiales—, como su complemento, y está listo para su implementación inmediata. Es un "terso texto de sólo 35 artículos", que refiere a los demás Códigos cuando lo considera apropiado, pero para la mayoría de los propósitos puede funcionar por sí mismo. Propone un número de mejoras futuras, que dependen de que se hagan reformas en los Códigos Especiales para poder efectuarse "y que esperablemente disparen esas reformas". Una de ellas es el requerimiento de registrar nombres nuevos y actos nomenclaturales (por ejemplo las designaciones de lectotipos), que en la nueva versión fue modificada para tomar ventaja de los recientes progresos tecnológicos. Otras innovaciones propuestas son el requerimiento de que las descripciones de nuevos taxones sean en inglés o latín (en el 2011 todavía se debían hacer en latín para plantas, y en cualquier lengua en animales), la opción de proteger nombres y sus atributos por medio de Listas de Nombres Adoptados, el prospecto de evitar los homónimos entre-Códigos para los nombres futuros, y el permiso, para los llamados organismos "ambirregnos", de usar sufijos de cualquier Código para los nombres supragenéricos, independientemente del Código que se esté aplicando. Introduce rangos adicionales a los primarios (profamilia entre familia y subfamilia, progénero entre género y subgénero, proespecie entre especie y subespecie) de forma de implementar las reglas de forma retroactiva para todos los tipos de organismos sin efectos desestabilizadores en el sistema actual de nombres. (W Greuter, secretario de la ICB[369]).
A comienzos de este siglo, la rápida evolución de las bases de datos dejó en evidencia que es más práctica la producción de listas de nombres adoptados por taxón, y la nueva edición fue creada para que sus provisiones puedan ser adoptadas en diferente momento por diferentes taxones dentro de cada Código, lo cual estaría determinado por el cuerpo internacional mandatario apropiado para cada taxón si fuera operacional (DL Hawksworth).[151]
Algunos cambios menores fueron realizados en los Códigos Especiales desde 1997[151][370] Con la llegada de los análisis moleculares aumentó la cantidad de grupos que fueron tratados sucesivamente bajo los Códigos de Botánica y Zoología, por lo que se acentuó la necesidad de armonizar la nomenclatura de protistas, que al 2011 queda pendiente. También queda pendiente un registro mandatario de nombres y una lista de nombres en uso, que al 2011 sólo sigue existiendo en bacterias (DL Hawksworth).[151]
Las variaciones de terminología entre Códigos, y la terminología unificada propuesta:
BioCode (unificado) | BC (de Bacterias) | ICBN (de Botánica) | ICNCP (de Plantas Cultivadas) | ICZN (de Zoología) |
|---|---|---|---|---|
Publicación y fecha de los nombres | ||||
| publicado | efectivamente publicado | efectivamente publicado | publicado | publicado |
| fecha | fecha | fecha (o prioridad) | fecha | prioridad |
| precedencia | prioridad | prioridad | precedencia | precedencia |
| anterior | senior | anterior | anterior | senior |
| posterior | junior | posterior | posterior | junior |
Estatus nomenclatural | ||||
| establecido | válidamente publicado | válidamente publicado | establecido | disponible |
| registro | validación | registro | registro | ------ |
| aceptable | legítimo | legítimo | ----- | potencialmente válido |
Estatus taxonómico | ||||
| aceptado | correcto | correcto | aceptado | válido |
Tipos o especímenes de referencia | ||||
| tipo portador de nombre | tipo nomenclatural | tipo nomenclatural | estándar nomenclatural (nombre del espécimen o fotos de referencia; ya que el "tipo" es toda la descripción original, que es invariable)[nota 17] | tipo nomenclatural |
| taxón nominal | nombre y tipo | nombre y tipo | ------ | taxón nominal |
Sinonimia | ||||
| homotípica | objetiva | nomenclatural u homotípica | ------ | objetiva |
| heterotípica | subjetiva | taxonómica o heterotípica | ------- | subjetiva |
| nombre de reemplazo (replacement name) | ------- | sustituto declarado (avowed substitute) o nombre de reemplazo (replacement name) | ------- | reemplazo explícito (explicit replacement) |
Prescindiendo de las reglas | ||||
| conservación | conservación | conservación | ------- | conservación |
| supresión | rechazo | supresión explícita | ------- | supresión |
Propuestas de un registro central de nombres
Para cerrar las "listas de nombres adoptados"[151] Charles Godfray (2002[229]) propuso una "primera revisión a través de la web" como el nuevo punto de partida de la nomenclatura en cada grupo, de forma que luego de una exposición por un tiempo predeterminado de las listas provisorias con su revisión taxonómica abiertas a los agregados de información nueva por parte de la comunidad con acceso a internet, éstas fueran cerradas y las descripciones previas encontradas posteriormente fueran consideradas como descripciones con nombres informales. Propuestas como ésta suelen estar acompañadas por la propuesta de registrar los nuevos actos nomenclaturales en alguna database central, de forma que el nombre no fuera establecido si no fue registrado. Alessandro Minelli (2003[253]), el presidente de la Comisión Internacional para la Nomenclatura Zoológica,[138] también argumentó sobre lo prioritario de contar con un registro central de nombres. El Código de Bacterias ya tiene un registro central de nombres: todos los nombres establecidos de las bacterias ahora son publicados en una única revista, con una nueva fecha de comienzo de la nomenclatura fijada en 1980.[371] Se esperaban reformas similares en los Códigos de macroorganismos, pero zoólogos y botánicos todavía tienen que ratificar las propuestas.[138] La posibilidad de que unos pocos especialistas pudieran dejar publicaciones fuera de la indexación y monopolizar la nomenclatura de un grupo había sido una de las preocupaciones que llevó a los botánicos a rechazar las llamadas para un registro central de nombres en el Congreso Internacional de Botánica en 1999, propuestas similares fueron rechazadas en el Congreso de Zoología de 1999.[138]
La posible reglamentación que había sido parte del borrador del BioCode 1997, fue modificada en el borrador del BioCode 2011 para incorporar las nuevas tecnologías y la posibilidad de dividir los taxones abarcados por los Códigos en grupos de tamaño más pequeño, cada uno con su propio cuerpo mandatario internacional.[151] En el año 2012 una enmienda[372] del Código de Zoología oficializó un registro central de nombres llamado Zoobank,[373] la enmienda no modifica sin embargo la fecha de inicio de la nomenclatura de cada grupo, ni el registro es mandatario, sólo insta a que a partir del momento se informen las publicaciones taxonómicas en esa web? En Botánica sólo el Mycobank, que había sido creado en el 2004, se adoptó como registro mandatario desde el 2013.[374] Al 2015 las listas no están cerradas.
El PhyloCode
Algunos autores argumentan problemas usando el sistema linneano de clasificación cuando se lo basa en el árbol filogenético (p. ej. de Queiroz y Gauthier 1990).[93] El PhyloCode (http://www.ohio.edu/phylocode),[217] creado en 1998 y actualizado periódicamente, está basado en lo que sus autores llaman una "aproximación filogenética" a la nomenclatura. Con el objetivo de ser un código de nomenclatura, tiene "varias similitudes en sus metas y en sus métodos" con lo que los autores llaman la "nomenclatura basada en rangos" (lo que otros autores llaman la nomenclatura linneana): provee métodos inambiguos para aplicar nombres a los taxones, para seleccionar un único nombre aceptado para cada taxón, y no impide a los taxónomos aplicarles rangos taxonómicos, "contrariamente a un concepto erróneo frecuente". Una de las diferencias es que el PhyloCode aplica los nombres a "la secuencia de ancestros y descendientes, o sus productos, los clados"; en lugar de aplicarlos "a una circunscripción + rango". De esta forma el Código define un taxón nominal[cita 68] cuando realiza la asociación entre un nombre y un clado,[93] o más rigurosamente, entre un nombre y las hipótesis cladísticas delimitadas por los especificadores (otros taxones o especímenes de referencia y apomorfías) de un clado, como publicadas en la primera descripción; lo que los autores llaman las "definiciones del nombre".[cita 68] Tres tipos de "definiciones del nombre" son la basada en nodos, la basada en ramas, y la basada en apomorfías. Una definición del nombre Asteraceae podría ser "el clado que se deriva del ancestro común más reciente de Barnadesia y Aster".[cita 68] Los nombres de los taxones no cambiarían con los cambios de circunscripción mientras se mantenga ese tipo, y como esos nombres no dependen del rango taxonómico (como en la nomenclatura de los Códigos que tiene reglas relacionadas al respecto, como el uso de sufijos), entonces cuando hay cambios de categorías taxonómicas no es necesario que cambien los nombres de los taxones (de Queiroz 2012).[28]
Los autores del PhyloCode pretenden reemplazar los Códigos de la nomenclatura linneana con su nuevo Código, lo cual no llegó a discutirse en el debate acerca de una taxonomía unitaria en el ambiente científico.[138][376]
Sin embargo, el PhyloCode ha sido intensamente debatido en revistas científicas de prestigio como Taxon,[26][377] y hasta aparecieron remeras universitarias con inscripciones "anti-rangos" que fueron sospechadas de propaganda hacia ese Código.[106][10] La crítica principal hacia el PhyloCode es que el sistema lineano es igual de compatible con el árbol filogenético, por lo que no hay necesidad de cambiar todo un sistema nomenclatural por otro.[377][10] De hecho el PhyloCode puede no ser compatible con el árbol filogenético en la categoría especie, ya que no maneja todos los conceptos taxonómicos utilizados en esa categoría.[377] Otra crítica es que ese "tipo" no probó ser en realidad más estable, "a menos que se redefina estabilidad",[377][10] por lo que aun dentro de una "filosofía taxonómica" cladista sería difícil evitar la convivencia con otros Códigos especialmente en categorías muy altas.[377] Quitar las categorías absolutas quitaría información sobre exclusiones y la mayoría de los taxónomos ve como suficientemente útil para la comunicación que estén impresas en los sufijos como para abandonarlos en las categorías del grupo familia.[10][106] (pero ver[86]) La posibilidad de que más de un espécimen de referencia, y las apomorfías o la descripción de un epíteto descriptivo puedan agregarse formalmente al "tipo" —lo último de hecho mandatario en taxones de categorías más altas—,[nota 18] y otras propuestas se podrían hacer bajo los Códigos en uso (p.ej.[377],[106]), que serían aprobadas después de un debate adecuado y un amplio consenso. Los "nombres descriptivos" de categorías altas de hecho cuando hacen referencia a un carácter o un plan corporal sobresaliente se interpretan como derivados de hipótesis cladísticas que contienen las apomorfías que definieron el plan corporal en su linaje ancestral y que le dan el nombre al grupo —que no necesariamente es monofilético—. Por ejemplo Cavalier-Smith (2013)[378] define al reino Fungi como derivado del linaje que adquirió rizoides absorbentes, tempranamente acoplado en su desarrollo a la adquisición de una pared celular de quitina con pérdida de la fagocitosis, propone a Excavata como grupo parafilético, y propone abandonar el nombre del supergrupo Unikonta ("un cilio"), ya que se había determinado que el cilio se había originado dos veces independientemente dentro del grupo.[378] Del aluvión de publicaciones referentes al PhyloCode, las dos lecturas recomendadas por el FAQ del ICZN son Wheeler (2004[24]) y Knapp et al. (2004).[156]
 Los nombres de los supergrupos de eucariotas ubicados en el árbol según la filogenia de Cavalier-Smith (2013[378]). |
El PhyloCode fue sólo una de las manifestaciones de los taxónomos que, en general bajo la nomenclatura de los Códigos en uso, señalan que es buena práctica que sea explicitado el "concepto de taxón" utilizado (p.ej.[117],[137],[122],[132] para los conceptos de especie,[263]en categorías intermedias). En taxones intermedios, un ejemplo del 2007 en subfamilias de rosáceas ha quedado así:[379]
| Subfamily Dryadoideae Juel, Kongl. Svenska Vetensk. Akad. Handl. n.s. 58: 55 (1918): | Subfamilia Dryadoideae Juel, Kongl. Svenska Vetensk. Akad. Handl. n.s. 58: 55 (1918): |
| The most inclusive clade containing Dryas (as typified by Dryas octopetala L.), but not Rosoideae (cf. above) nor Spiraeoideae (cf. below). | El clado más inclusivo conteniendo a Dryas (como tipificado por Dryas octopetala L.), pero no Rosoideae (cf. más arriba) ni Spiraeoideae (cf. más abajo). |
| Included taxa: Cercocarpus H. B. and K., Chamaebatia Benth., Dryas L., Purshia DC. (including Cowania D. Don) | Taxones incluidos: Cercocarpus H. B. and K., Chamaebatia Benth., Dryas L., Purshia DC. (incluyendo Cowania D. Don) |
| Distinctive non-molecular features: Shrublets, shrubs, or small trees. Symbiotic nitrogen fixation present. Cyanogenic glycosides present. Sorbitol present in trace amounts. Leaves compound in Chamaebatia, simple in the other genera. Stipules present. Hypanthium free from ovary. Pistils 1 (Cercocarpus, Chamaebatia, Purshia) or 4-many (Cowania, Dryas). Fruit an achene or achenetum. Base chromosome number x=9. | Características distintivas no moleculares: Arbustillos, arbustos, o arbolillos. Fijación de nitrógeno simbiótica presente. Glucósidos cianogenéticos presentes. Sorbitol presente en cantidades traza. Hojas compuestas en Chamaebatia, simples en los demás géneros. Estípulas presentes. Hipanto libre del ovario. Pistilos 1 (Cercocarpus, Chamaebatia, Purshia) o 4-muchos (Cowania, Dryas). Fruto aquenio o aquenetum. Número de cromosomas básico x=9. |
Potter et al. 2007[379] | |
La diferencia entre ambos Códigos está en el tipo, que se traspasa de forma inalterable de taxón a taxón.
Véase también
Este artículo trató sobre el campo de acción de la taxonomía. Para la clasificación de la biodiversidad ver:
Clasificación de los organismos en categorías altas (reinos, dominios, imperios) trata los conceptos de macroevolución que modelan la clasificación, las diferencias entre clasificaciones cladistas y evolucionistas, y la cantidad de reinos y cómo fueron madurando hasta la actualidad.
Las taxonomías seleccionadas para clasificar las plantas cultivadas en:
- Cultivar
- Clasificación y nomenclatura de plantas cultivadas
Otras ciencias relacionadas:
Sistemática que abarca a la taxonomía y al análisis filogenético.
Los códigos de nomenclatura:
Código de Nomenclatura una lista de los códigos en uso.
- Código Internacional de Nomenclatura Zoológica
- Código Internacional de Nomenclatura Botánica
- Código Internacional de Nomenclatura de Bacterias
- Código Internacional de Nomenclatura para Plantas Cultivadas
- BioCode
- PhyloCode
Otros artículos relacionados:
- Taxón
- Concepto de Taxón
- Circunscripción (taxonomía)
Escuelas de la Sistemática
- Escuela cladista
- Escuela fenética
- Escuela evolucionista
- Sistema natural de clasificación
- Sistema artificial de clasificación
- Taxonomía popular
- Categoría taxonómica
- Autor de nombre científico
Notas
↑ ab Definición de dendrograma en libros de matemática como por ejemplo en Everitt et al. 2011[1] Un dendrograma es un árbol donde cada nodo representa la agrupación de todos los nodos terminales conectados a él. La "topología del árbol" está dada exclusivamente por las relaciones entre nodos. Un dendrograma es una representación pictórica de un proceso de agrupamiento jerárquico, sea éste aglomerativo (como el de la figura) o divisivo, y es la aproximación a la filogenia o a la clasificación que utilizan todas las escuelas de taxonomía.

Dendrograma aglomerativo
(Sobre los dendrogramas) "Como el sistema lineano requiere que las entidades taxonómicas sean ordenadas de una forma jerárquica y sin solapamientos, la mayoría de los métodos de agrupamiento usados en taxonomía biológica han sido técnicas jerárquicas, sin solapamientos."
Sokal (1986[2])
Ver también cita "clado".[cita 1]
↑ Los libros de texto aparentemente la suelen llamar "jerarquía relativa" o "rangos relativos", debido a que están jerarquizados entre ellos. En este texto, por "relativo" se entiende que el rango cambia según el taxón que se use como referencia (p.ej. son rangos relativos "abuelo", "padre", "nieto").
↑ ab Se puede encontrar la diferencia entre la definición de "escuela fenética" y la de "taxonomía numérica" por uno de los padres de la escuela fenética: Sneath, P. H. (1995[67]), quien define a la fenética como la filosofía y a la taxonomía numérica como los métodos de análisis cuantitativos —hoy en día por computadora—, y afirma que así las habían concebido en la publicación original de Sokal y Sneath (1963[47]).
↑ Esta afirmación es una respuesta común a los autores del PhyloCode, quienes pretenden estabilidad en la nomenclatura proponiendo que los nombres se apliquen al clado real o "entidad concreta" (de Queiroz y Gauthier 1990[93]).
↑ El término "taxonomía integrativa" hace referencia a la práctica habitual y deseada en biología sistemática, de tomar datos de líneas de evidencia diferentes, y encontrarles congruencia en la delimitación de los taxones mediante un marco teórico evolutivo y biológico. El término en sí sin embargo es nuevo, fue acuñado independientemente y casi al mismo tiempo por Dayrat (2005[99]) y por Will et al. (2005[100]), los dos en respuesta a los altamente publicitados en esa época análisis moleculares de ADN y "ADN barcodes" como únicas fuentes de evidencia en la delimitación e identificación de las especies respectivamente.
↑ Hennig proponía que la edad geológica absoluta de un grupo fuera el criterio para decidir su categoría taxonómica, y hay científicos trabajando para lograr esta meta en algunos grupos, como en Nevo y Wasser (2004[103]). Sin embargo Wiley y Lieberman (2011[104]) mencionan que la edad cronológica absoluta para decidir la categoría taxonómica es un concepto abandonado.
↑ Para la escuela cladista o al menos para Judd et al. (2007[18]), las categorías taxonómicas por arriba de la especie son arbitrarias, debido a que "un género (grupo de especies) en una familia puede no tener la misma edad ni albergar la misma cantidad de variación, ni de hecho tener nada en común con un género de otra familia, más que el hecho de que los dos son grupos monofiléticos que pertenecen a la misma categoría taxonómica" (esta definición difiere de la definición de categoría taxonómica evolucionista). Ésta es una de las disputas entre las dos escuelas, ya que en las clasificaciones cladistas en busca de grupos monofiléticos, como resultado los géneros, las familias, etcétera no son unidades comparables (Stevens 1997[108] citado en Judd et al. 2007[18]). Esta diferencia entre las dos clasificaciones afecta otras ramas de la biología que las utilizan como herramienta. Por ejemplo, "es común ver medidas de diversidad como un listado de las familias o géneros en un lugar dado", pero si la clasificación es cladista, "el hecho de que esos taxones pertenezcan a una "familia" no significa nada en particular" (Judd et al. 2007[18]).
↑ Algunos proponentes del PhyloCode, pero no todos, proponen eliminar las categorías taxonómicas. Para una distinción ver De Queiroz (2006[109])
↑ Sokal (1986[2]) define nicho de forma más laxa, de forma que cada categoría taxonómica pueda ser reconocida por un "nicho" cada vez menos estricto. Pero nótese que por arriba de la categoría especie no hay forma de definir un taxón de forma de definir la sobrevivencia de sus linajes. Mayr (1981[4]) prefiere el término "zonas adaptativas" (adaptive zones) para referirse a esos espacios multidimensionales. Véase también cita a Nixon y Carpenter (2000[106]) en [cita 35]
↑ Además de esos tres Códigos (Zoología[31] Botánica,[31] Bacterias[30]) hay códigos internacionales de nomenclatura de otras disciplinas más pequeñas: el de de plantas cultivadas[141], el de bacterias patógenas de plantas[142], y el de virus[143]. Estos códigos están avalados por ejemplo por el IUBS, autores también del BioCode.[33] Los protistas se nombran bajo el código de botánica o el de zoología, según en qué disciplina se haya considerado históricamente a cada uno de sus grupos.
↑ En este texto una definición por extensión, se refiere a qué organismos componen el taxón. Se puede encontrar en la literatura taxonómica el término "relaciones extensionales" o "relacionados por extensión", que por claridad en este texto serán evitadas, pero que se pueden encontrar por ejemplo en el review de Dupuis (1984[41] ), traducción:
"Cualquier aproximación a la clasificación (classificatory approach) usualmente considera "objetos" (objects) —aquello que está siendo clasificado, por ejemplo, individuos biológicos o taxones— y sus "atributos" (attributes), muchas veces llamados "caracteres (characters) por los naturalistas. Las relaciones entre objetos y atributos (en biología, entre taxones y caracteres) pueden ser vistas como relaciones por intensión (intensional), como si los atributos determinaran objetos (as if atrributes make objects), o como relaciones por extensión (extensional), como si los objetos determinaran los atributos, ver debajo. De acuerdo con una visión por intensión, la clasificación se basa en la similitud de atributos; de acuerdo a la visión por extensión, depende de posibles relaciones intrínsicas entre los objetos. Entre los objetos biológicos –individuos y taxones– que están enlazados por una historia, estas relaciones hacen posible una visión por extensión que es precisamente la expresada por la declaración fundamental Hennigiana (the Hennigian fundamental statement)."
↑ La nomenclatura de bacterias no es independiente de protozoos, algas y hongos, como se lee en el Código de Bacterias (Revisión 1990[30]): Principle 2. The nomenclature of bacteria is independent of botanical nomenclature, except for algae and fungi, and of zoological nomenclature, except protozoa. For these exceptions and for relationships with virological nomenclature, see General Consideration 5 and Rule 51b(4). "Independent" means that the same name may be validly used for a taxon of bacteria as well as a taxon of plants or animals with the exceptions noted above.
↑ ab Técnicamente, los Códigos Internacionales de Nomenclatura Zoológica y Botánica especifican que los nombres de los géneros y de las especies deben ir en una tipografía diferente de la del resto del texto. Normalmente eso es en itálicas en textos tipeados y subrayado en textos escritos a mano, pero no necesariamente tiene que ser así. Además y por esa razón, en textos tipeados que están enteramente en itálicas, los nombres de géneros y especies pueden estar escritos en cualquier tipografía menos itálicas, por ejemplo pueden ir en letras redondas, o pueden ir en itálicas si además están subrayados.
Citas sobre el uso de itálicas en los Códigos de Zoología, Botánica y Bacterias:- Código Internacional de Nomenclatura Zoológica[31], Recomendación General 6: "The scientific names of genus –or species-group taxa– should be printed in a type-face (font) different from that used in the text; such names are usually printed in italics, which should not be used for names of higher taxa. (…)"
- En el de Botánica[29] no hay artículo que lo mencione pero en el prefacio dice: "As in all recent editions, scientific names under the jurisdiction of the Code, irrespective of rank, are consistently printed in italic type. The Code sets no binding standard in this respect, as typography is a matter of editorial style and tradition, not of nomenclature. Nevertheless, editors and authors, in the interest of international uniformity, may wish to consider adhering to the practice exemplified by the Code, which has been well received in general and is followed in a number of botanical and mycological journals."
- En el de Bacterias[30] dice en la sección 4A, "Sugerencias para autores y publicadores": "For scientific names of taxa, conventions shall be used which are appropriate to the language of the country and to the relevant journal and publishing house concerned. These should preferably indicate scientific names by a different type face, e.g., italic, or by some other device to distinguish them from the rest of the text." http://www.ncbi.nlm.nih.gov/books/NBK8816/
- Código Internacional de Nomenclatura Zoológica[31], Recomendación General 6: "The scientific names of genus –or species-group taxa– should be printed in a type-face (font) different from that used in the text; such names are usually printed in italics, which should not be used for names of higher taxa. (…)"
↑ A diferencia de lo que sucede en nombres de géneros y especies en los que hay consenso en los Códigos y cuyo uso es seguido por todas las editoriales sin excepción, con respecto a la tipografía de los taxones superiores a género hay diferentes opiniones, si bien las recomendaciones sobre su uso se encuentran en las mismas secciones de los Códigos que mencionan que los géneros y especies deberían ir en itálicas: el de Zoología "recomienda" las redondas por arriba de género, mientras que el de Botánica en las últimas ediciones y el de Bacterias lo dejan a criterio de cada editorial pero "el que ellos mismos usan" (Botánica) o "sugieren" (Bacterias) son las itálicas.[nota 13] El uso más común en las editoriales científicas, tanto de Zoología como de Botánica, es el de las redondas por arriba de género. En Botánica, incluso en la misma revista Taxon que es donde se publican las propuestas sobre cambios en el Código de Botánica o pedidos sobre decisiones de cómo aplicarlo (además de otros trabajos nomenclaturales y de sistemática y evolución)[cita 51], se usan redondas por arriba de género salvo en trabajos de índole estrictamente nomenclatural, como se puede leer en sus "Indicaciones para autores" todavía en el 2013.[cita 52] Por lo que la convención por ahora, en trabajos no estrictamente nomenclaturales, siguen siendo las redondas por arriba de género, convención que puede cambiar en el futuro.
↑ Los antiguos griegos veían a la especie humana como una especie de animales, como quedó documentado en la taxonomía de Aristóteles plasmada en su Historia animalium. Sin embargo en la Edad Media europea se quitó a los hombres de la taxonomía animal argumentando que tenían alma. En el siglo XVI comenzó la tendencia a incluirlos de nuevo entre los animales, probablemente bajo la influencia de la incipiente ciencia de la anatomía animal comparada.[207]
↑ Institute of Scientific Information o ISI, parte de Thomson Reuters. El factor de impacto de ISI es la medida más antigua y una autoridad en mediciones de prestigio, si bien tiene sus falencias y puede ser manipulada. ISI es desde principios de los 1990 parte de Thomson Reuters. El factor de impacto IF creado por ISI no fue originalmente creado con los fines con los que se lo utiliza hoy en día, la historia de ese cambio en Boero (2010[249]). Descripción y comentarios en Steele et al. (2006[227]).
↑ Las reglas para construir un “estándar nomenclatural” que puede ser un ejemplar o fotografías están en el Código Internacional de Plantas Cultivadas (Brickell et al. 2009[215]) en la "DIVISION V. NOMENCLATURAL STANDARDS.", véase también el "APPENDIX III. PLACES MAINTAINING NOMENCLATURAL STANDARDS".
↑ Un ejemplo con la posibilidad de que un epíteto descriptivo se agregara formalmente al "tipo" (p.ej.[377],[106]), como hasta ahora sólo está reglamentado en categorías superiores al grupo familia (u orden en bacterias). En 1930 Pangalo había separado de las calabazas de Cucurbita moschata un grupo de hibridación algo parecido morfológicamente, al que llamó Cucurbita mixta. Posteriormente debió corregirse el nombre a Cucurbita argyrosperma ("Cucurbita de semillas plateadas"), la hoy primera variedad descripta formalmente con nombre binomial. La especie biológica en la que se incluye normalmente no posee semillas plateadas. Si el epíteto hubiera conservado su nombre descriptivo hoy la especie se llamaría Cucurbita mixta y ese epíteto se utilizaría para referir a la variedad de Cucurbita mixta con semillas plateadas cuyo nombre vulgar es Silver Seed Gourd ("calabazas de semillas plateadas"). Un ejemplo en la categoría de familia podrían ser las aráceas, en las que la taxonomía cladista incluyó formalmente a las lemnáceas (las lentejas de agua). Si las aráceas hubieran conservado su relación con la descripción de familia "aroide" se hubiera descartado el nombre y creado otro nuevo para la familia que incluye aráceas sensu stricto + lemnáceas. Es lo que sucede en categorías de rangos más altos. En la época en que las especies eran los actuales ecotipos, Linneo tuvo el poco tacto de nombrar con el epíteto sapiens a la raza europea. Al corregir la circunscripción del epíteto con el agregado de información nueva, el epíteto sapiens hubiera quedado para nombrar toda la especie, y el ecotipo al que pertenecía Linneo hubiera debido recibir otro epíteto nuevo.
Citas
↑ abcdefgh En la argumentación original de Hennig (1950[36] en alemán, 2ªed 1966[37] en inglés) –considerado el fundador del cladismo, o "su voz más articulada" como lo llamaba Mayr 1982[38] p.167.–, y como se puede ver por ejemplo en el cuadro "Hennigian Argumentation" de Judd et al. (2007[18] p. 20.), son las apomorfías las que determinan la topología del árbol y por lo tanto sus secuencias de ancestros y descendientes, asumiendo que cada vez que se generó una línea filética hubo cambios evolutivos (apomorfías) en ella; si bien también se pueden usar otros métodos para obtener la topología del árbol. Autores posteriores se refirieron a los árboles de Hennig como cladogramas y a sus ramificaciones derivadas de un corte como clados –el primero en utilizar el término "cladismo" en reemplazo del confuso término Hennigiano "sistemática filogenética" fue Mayr (1965[39])–, pero además se utiliza "clado" en un sentido más amplio, como hipótesis de ser todos los derivados de una secuencia de ancestros y descendientes con un ancestro común. Muchos libros de texto incluyen los ancestros en el grupo, p. ej. Judd et al. (2007[18])p. 5-6., traducción: "Los grupos monofiléticos también son referidos como clados", "Una aproximación filogenética demanda que cada taxón sea un grupo monofilético, definido como un grupo compuesto por un ancestro y todos sus descendientes –mono: único, phylum: linaje)"–. Esta definición resulta poco rigurosa ya que los métodos de obtención de clados lo hacen sobre los organismos de los que se tomaron los datos: las poblaciones vivientes. Una de las voces más conspicuas del cladismo como K de Queiroz –uno de los creadores del PhyloCode– reconoce en su publicación del 2012[28] que su intención es nombrar a la ...secuencia de ancestros y descendientes, o sus productos, los clados. De la misma forma Cavalier-Smith (2010[40]), llama "clado Hennigiano" al dendrograma formado por el método de las apomorfías: "Uno no podría enfatizar lo suficientemente fuertemente que un cladograma Hennigiano no es un árbol filogenético, los cladogramas no tienen ancestros, sólo especies vivientes". El término monofilético también fue utilizado sin los ancestros por GG Simpson y E Mayr y al menos según Mayr fue tradicionalmente definido así, como un adjetivo que califica a un taxón —a una circunscripción—, como se puede leer en las citas textuales de "monofilético sensu Hennig".[cita 15]
Sobre la etimología de clado, así lo cuenta Dupuis (1984[41]), traducción: "Cuando acuñó el término "clado" (clade) a partir del término "cladogénesis" (cladogenesis)... que fue tomado directamente de Rensch,[42] "Huxley intentó denotar "unidades monofiléticas delimitables" (Huxley 1957[43] p.454) o, más precisamente, "unidades monofiléticas de cualquier magnitud" (Huxley 1958[44] p.27). Él parece no haber conocido un uso más antiguo de este término en Cuénot 1940[45] (...(sin la misma definición)...). El adjetivo "cladístico" (cladistic) ha sido explícitamente derivado de "clado" (clade) sensu Huxley por Cain y Harrison, para quienes "la cercanía de las relaciones genealógicas (relationship) en términos de líneas filéticas pueden ser llamadas cladísticas..., las similitudes debido a un ancestro común, no a una convergencia, pueden ser llamadas patrísticas" (Cain y Harrison 1960[46] p.3). Tanto cladístico como patrístico (Cain y Harrison 1960[46]) refieren a afinidades filéticas y necesitan la clarificación que fue abordada por Sokal y Sneath (Sokal y Sneath 1963[47]:220): Cladística es una relación "por el ancestro común más reciente" (through recency of common ancestry), y patrística, una por ancestro común sin más especificaciones. Para esos autores, que confían en las relaciones (relationships) de los tipos opuestos de Hennig I y Ia (Hennig 1957[48] fig.6), cladística es "el tipo I de las relaciones filogenéticas (phylogenetic relationship) de Hennig 1957[48]". Entonces igualan una relación cladística (cladistic) con una Hennigiana. El término "cladista" (cladist) fue introducido en 1965 "en línea con la terminología desarrollada por Rensch (1960), Huxley (1958[44]) y Cain y Harrison (1960[46])" (Mayr 1965a[49]:167). Si bien ninguno de esos autores se refería a Hennig, al acuñar el término "cladismo" (cladism) Mayr tipifica "al cladista" (the cladist) como a quien adhiere a aquella "escuela filogenética" de la cual Hennig es el "vocero más articulado" (Mayr 1982[38]:167). El término "cladograma" (cladogram) fue introducido en ese mismo año (1965) pero tiene dos significados diferentes. Para Mayr (1965b[39]:81), "el cladograma de los cladistas" es el diagrama filético de la escuela Hennigiana. Para Camin y Sokal (Camin y Sokal 1965[50]:312), el término "cladograma" es usado "para distinguir un dendrograma cladístico de uno fenético que podría ser llamado un fenograma", desafortunadamente, estos autores construyeron sus "cladogramas" por técnicas fenéticas y no Hennigianas. Consecuentemente, hay dos tipos de cladogramas: aquellos que se basan en atributos (el Stufenreihe de Hennig, los sinapomorfogramas —synapomorphograms— de Sneath, ver Moss 1983[51]) y aquellos que se basan en objetos (es decir en construcciones fenéticas); el utilizarlos inadvertidamente como sinónimos (por ejemplo en Sokal 1983[52]:160) lleva a confusiones. En los trabajos de Hennig, estos términos son siempre utilizados entre comillas y aparecen como meras citas. En trabajos de otros autores, "cladista" y "cladismo" comúnmente aplican hoy en día a la escuela nacida por el pensamiento Hennigiano, mientras que "cladístico" y "cladograma" se han vuelto igualmente populares tanto en la escuela fenética como en la Hennigiana, y consecuentemente, su significado se mantiene ambiguo. Aún peor, "cladograma" se ha vuelto una palabra de moda que es a veces utilizada para denotar construcciones intuitivas que no hacen referencia a un procedimiento definido."
Ver también la nota "dendrograma".[nota 1]
↑ abcd :
Como Hull (1979[3]) ya ha hecho notar, los cladistas en realidad clasifican caracteres en lugar de organismos, tienen que hacer la suposición arbitraria de que nuevas apomorfías se originan cada vez que una línea [filética] se ramifica de su línea hermana. Esto es poco probable en la mayoría de los casos.
E Mayr (1981[4])
Las clasificaciones cladistas no representan el orden de ramificación de grupos hermanos, sino el orden de emergencia de caracteres derivados únicos.
DL Hull (1979[3]) citado en E Mayr (1981[4]:nota 28).
Nosotros utilizamos el cladograma en el sentido de un sumario jerárquico de los datos de caracteres, no contiene necesariamente supuestos sobre, ni hace predicciones de, evolución y especiación.
Nelson y Platnick (1981[5]:14), un libro muy utilizado en su época.
Los sistemáticos siempre han sido, son, serán, y deberían ser tipologistas.
Nelson y Platnick (1981[5]:328)
Esta conceptualización ha llevado a que G Nelson, NI Platnick y también C Patterson (por obras como C Patterson 1982[6]), fueran llamados pattern cladists ("cladistas de patrones") por Beatty (1982[7]), Brady (1982[8]) y otros de la época (Platnick lo llamó transformed cladistics en 1979[9]).
El concepto Hennigiano [de especie], que data desde los 1960's, es definitivamente una mejora, la visualización de las especies como los entrenudos [de un árbol filogenético] --aquellas unidades que se desprenden de eventos de especiación sucesivos (o en algunos casos un evento de especiación más un evento de extinción posterior) [...] De acuerdo con esta mirada, sin embargo, las especies del tallo ancestral no pueden sobrevivir los eventos de especiación, éstas cesan de existir aun si una de sus especies descendientes resulta ser idéntica en todas las características por las que se la conoce. Cualquiera de estos ejemplos claramente representaría un caso de triunfo de la teoría sobre los datos. [...] Los taxones superiores putativos, bajo el concepto de especie filogenética real, deben ser evidenciados por caracteres autapomórficos, pero las especies no tienen como requisito la aparición de una autapomorfía, sólo necesitan una combinación de caracteres única. En otras palabras, las especies deben ser diagnosticables, no necesariamente autapomórficas.
Platnick (2001[10])
Sobre la relación entre "clado" y "monofilético":
Los cladogramas son esquemas de clasificación que testean la monofilia. Discusiones recientes argumentaron que los cladogramas ignoran las genealogías en la clasificación. La conclusión de estas discusiones es que hay dos formas de interpretar una clasificación: sincronía (synchrony) y diacronía (diachrony).
Ebach y Williams (2004[11])
...Recientemente, Podani (2010[12]) trató de resolver el problema de los diferentes rangos temporales (time levels) restringiendo los términos monofilia y parafilia a los áboles filogenéticos (que son diacrónicos, es decir que cada linaje en realidad se mantiene a través de varios rangos temporales), mientras que los términos monocladístico y paracladístico (monoclady and paraclady) se usarían para describir cladogramas (que son sincrónicos y por lo tanto confinados a un rango temporal) [...] La mayoría de los taxones en las clasificaciones cladísticas contemporáneas son vistos como monocladísticos, ya que derivan de clados, pero no son necesariamente monofiléticos.
[...]
Box 1, "Clado": Una rama de un cladograma, que representa una hipótesis de grupo holofilético (- monofilético s.str.).
Hörandl y Stuessy (2010[13])
El citado Podani (2010[12]) define grupos monofilético (rama del árbol de linajes), monocladístico (rama del cladograma), monotético (rama de un "cladograma de patrones", pattern-cladogram).
↑ Decía K de Queiroz (1992): "En 1969, el libro de Mayr, Principles of Systematic Zoology fue probablemente el mejor texto de sistemática y el mejor libro de referencia disponible (Mihener 1969), y retuvo un lugar prominente entre los de su estilo por años después de su publicación. Cuando fue publicado por primera vez, estaba actualizado, era considerado una autoridad en la materia, y era el más completo, y aún más de 20 años después de su apogeo continúo usando mi copia frecuentemente. Si bien muchos libros excelentes sobre el tema han sido publicados después, sólo el de Wiley (1981) Phylogenetics, sirve a tantos propósitos diferentes. Pero un montón ha sucedido desde 1969. Ningún libro sobre sistemática publicado hace tanto tiempo es actual hoy en día. Consecuentemente, la publicación de una edición revisada de Principles of Systematic Zoology se espera que sea un nuevo estándar en este campo."
↑ abc La definición de taxonomía que parece ser la más utilizada en los libros de texto (y la aquí utilizada) puede verse resumida en Simpson (2005[20]):
La sistemática está definida en este libro como una ciencia que incluye y abarca la taxonomía tradicional, la descripción, identificación, nomenclatura, y clasificación de los organismos, y que tiene como su meta principal la reconstrucción de la filogenia, o historia evolutiva, de la vida. Esta definición de sistemática no es nueva, pero tampoco es universal. Otros en el campo tratarían a la taxonomía y a la sistemática como áreas separadas pero superpuestas, aún otros arguyen que el uso histórico necesita lo que es en esencia una reversión de las definiciones utilizadas aquí. Pero las palabras, como los organismos, evolucionan. El uso de sistemática para describir un campo que abarque todos los aspectos es más útil y además representa el consenso de cómo la mayoría de los especialistas usa el término, siendo un ejemplo la revista Systematic Botany, que contiene artículos tanto de taxonomía tradicional como de reconstrucción filogenética."
Simpson (2005[20]):5
La definición concuerda con la de otros libros de texto, Judd et al. (2007[18]) es un libro cuya primera edición fue muy premiada:
La sistemática abarca la disciplina de la taxonomía, un término ligado a la palabra taxón.
Judd et al. 2007[18]:cap1:p6
Aunque el mismo Judd et al.luego en el capítulo 2, p. 13, las trata como sinónimos:
La biología sistemática (o taxonomía) es la teoría y práctica de agrupar individuos en especies, ordenar esas especies en grupos más grandes, y dar a esos grupos nombres, produciendo así una clasificación.
Judd et al. 2007[18]cap2:p13
La definición del respetado[18] Simpson (1961[15]):
"La taxonomía es el estudio teórico de la clasificación, incluyendo sus bases, principios, procedimientos y reglas".
GG Simpson (1961[15]:11) como citado en QD Wheeler (2008[21]:5)
Ver también la cita "Definición de sistemática".[cita 5].
↑ ab
El autor E Mayr (1969[16]) en Principles of Systematic Zoology sigue a GG Simpson[15] en la distinción entre taxonomía (la teoría y práctica de clasificar organismos) de la sistemática (la ciencia de la diversidad de organismos). Es ventajoso que uno de estos dos términos no se desperdicie volviéndolos, como en el pasado reciente, sinónimos, si bien uno puede argumentar cuál es cuál.
Richards (1970[17]), review del libro de E Mayr (1969) Principles of Systematic Zoology.
Al 2007 la primera cita del libro de texto de los cladistas Judd et al. (2007[18]), y la única sobre este tema, es:
De acuerdo con el paleontólogo George Gaylord Simpson (1961[15]:7), "la sistemática es el estudio científico de los tipos (the kinds) y la diversidad de organismos y cada una y todas las relaciones entre ellos".
Judd et al. (2007[18]), un libro cladista cuya primera edición en 1998 fue muy premiada.
Ver también la cita "Definición de taxonomía".[cita 4].
↑ ab
De acuerdo con el punto de vista del autor [E Mayr], que pienso que casi todos los biólogos deben compartir, la especie es la única categoría taxonómica que tiene al menos en los ejemplos más favorables una existencia completamente objetiva.
Richards (1970[17]), review del libro de E Mayr (1969) Principles of Systematic Zoology.
↑ abcdefgh Modernamente y deslindando las diferencias en las definiciones de Taxonomía —desde hace muchas décadas con una definición mínima— y Sistemática, el antigua área de la "Taxonomía Descriptiva" o "alfa-Taxonomía" es llamada más apropiadamente el área de la "Ciencia de la Biodiversidad", por ejemplo con esa definición explícita en la agencia de subsidios National Science Foundation de Estados Unidos:
Biodiversity: Discovery & Analysis program. Supports all aspects of Biodiversity Science, including expeditionary and exploratory research in natural environments to advance the discovery, identification, description, classification and cataloguing of the world's biodiversity. These investigations should be focused at the organismal level and within an evolutionary context. Proposals may target any group of organisms from any natural habitat. Proposals should address biodiversity discovery questions and may entail a primarily clade-based approach (i.e., regional to planetary species-level inventories for a particular taxonomic group), a guild-based approach (i.e., focused on ecological assemblages, common habitats, or host-symbiont assemblages), or a geographically-based approach. Such projects should focus on clades, guilds, or areas for which species-level biodiversity is very poorly known, and successful proposals will make a compelling case demonstrating that lack of knowledge. Projects that include fieldwork, work in existing collections, taxonomic identifications, revisionary classifications, monographs, hypothesis testing, and bioinformatics are appropriate in this program.
Programa de Biodiversidad: Descubrimiento y Análisis. Da apoyo a todos los aspectos de la Ciencia de la Biodiversidad, incluyendo la investigación expedicionaria y exploratoria en ambientes naturales para avanzar en el descubrimiento, la identificación, la descripción, la clasificación y la catalogación de la biodiversidad del planeta. Estas investigaciones deberían estar enfocadas en el nivel del organismo y dentro de un contexto evolutivo. Las propuestas pueden apuntar a cualquier grupo de organismos de cualquier hábitat natural. Las propuestas deberían abordar preguntas de descubrimiento de la biodiversidad y pueden implicar una aproximación primariamente basada en un clado (por ejemplo inventarios en el rango de especie regionales a planetarios para un grupo taxonómico particular), una aproximación basada en gremios (es decir, enfocados en asemblajes ecológicos, hábitats en común, o asemblajes hospedador-simbionte), o una aproximación basada en geografía. Estos proyectos deberían enfocarse en clados, gremios o áreas para las que la biodiversidad en la categoría de especie es muy pobremente conocida, y las propuestas exitosas serán muy persuasivas demostrando esa falta de conocimiento. Los proyectos que incluyen trabajo de campo, trabajo sobre las colecciones existentes, identificaciones taxonómicas, clasificaciones revisionarias, monografías, testeo de hipótesis, y bioinformática son apropiados en este programa".
Biodiversity: Discovery & Analysis proposals should include well-documented plans for fieldwork coordination and permitting, vouchering of new collections, specimen preparation, long-term specimen storage regimes that are openly accessible, specimen identifications and descriptions, georeferencing, data modeling and databasing, and rapid dissemination of data into public biodiversity databases. The rationale for targeting specific taxonomic groups/areas and proposed sampling approaches should be well-justified. Innovative approaches, such as rapid identification/description, cybertaxonomy, and automated identification tools, are encouraged.
Las propuestas de Biodiversidad: Descubrimiento y Análisis deberían incluir planes bien documentados de coordinación del trabajo de campo y permisos, depósitos de los especímenes de referencia de las colecciones nuevas, preparación de especímenes, regímenes de almacenamiento de especímenes de largo plazo que sean abiertos al público, identificación de especímenes y descripciones, georreferencias, modelado de datos y dababasing, y la diseminación rápida de datos en las bases de datos de biodiversidad públicas. Las razones por las que se apunta a grupos taxonómicos o áreas específicos y las aproximaciones al muestreo propuestas deberían estar bien justificadas. Las aproximaciones innovativas, como las de identificación/descripción rápida, la cibertaxonomía, y las herramientas de identificación automatizada, son alentadas.
NSF Systematics and Biodiversity Science Cluster (SBS) 1. Biodiversity: Discovery & Analysis program.[163]
↑
Sistemáticos, biólogos evolutivos, biólogos de poblaciones, biólogos de la conservación, ecólogos, agrónomos, horticultores, biogeógrafos y muchos otros científicos están más interesados en las especies que en ningún otro taxón.
Judd et al. (2007[18]):144
↑ ab "Resolución taxonómica" o precisión taxonómica como utilizados por Franz et al. (2008[22]).
↑ "Taxonomía linneana" y "jerarquías linneanas" suelen ser los nombres utilizados en referencia a esta taxonomía por autores como RR Sokal (1986[2]), DL Hull (1979[3]), Judd et al. (2007[18]:548: "...the use of Linnaean ranks..."), QD Wheeler (2008[21]:6) "...a formal Linnaean classification..." RK Brummitt[25] "...Linnaean classification" etc. Pero ver:
Stuessy (2000[26]:232):
“
-Comentario aparte, sería de ayuda si los defensores del PhyloCode dejaran de referirse al sistema nomenclatural botánico moderno como el "Sistema Linneano" (p. ej. Cantino 2000:87). Linneo delineó sus reglas de nomenclatura en Critica Botanica (1737), que tiene poca resemblanza con nuestro moderno Código Internacional de Nomenclatura Botánica (Greuter et al. 1994) desarrollado por mucha gente a través de muchas versiones a lo largo de los últimos 130 años.
”
de Queiroz (2000[27]) contesta refiriéndose a ella como "taxonomía tradicional" (en contraposición con el PhyloCode) y comenta (p.535-536)
de Queiroz (2000[27]):
“
- Stuessy también criticó a los defensores de la nomenclatura filogenética por referirse al actual sistema de nomenclatura botánica como el "sistema Linneano". Su razón fue que el moderno International Code of Botanical Nomenclature (Greuter et al. 2000) difiere en muchos aspectos de las reglas de nomenclatura delineadas en Linneo (1737) Critica Botanica. Si bien ciertamente hay diferencias entre el sistema nomenclatura descripto en el moderno código botánico y el sistema propuesto por Linneo, el sistema moderno es referido como "linneano" porque está basado en un método de definiciones similar al utilizado por Linneo, el cual es a su vez basado en las categorías de una taxonomía jerárquica que deriva largamente de su trabajo (ver de Queiroz 1997 para una discusión de la terminología). En este sentido, es apropiado referirse a este sistema de nomenclatura encarnado en el moderno Código Botánico como "un sistema linneano", si bien estoy de acuerdo con la crítica de Stuessy en reconocer que sería apropiado reservar el uso de "el sistema linneano" para hacer referencia al sistema utilizado por el mismo Linneo.
”
Más tarde (p.ej. de Queiroz 2012[28]) la llamaría:
de Queiroz (2012[28]):
“
-...Taxonomía basada en rangos (lo que otros autores llaman 'taxonomía linneana')...
”
↑ ab
En la visión de Aristóteles tres cosas pueden conocerse sobre una entidad: su esencia, su definición, y su nombre. El nombre nombra la esencia. La definición otorga una descripción completa y exhaustiva de la esencia. Se puede derivar que el nombre es el nombre de la entidad y la definición una descripción de la misma.
Hull (1965[32]:318)
↑ Definición de taxón según los Códigos en uso:
Código de Botánica (2012[29]), División I, Principio IV
Each taxonomic group with a particular circumscription, position, and rank can bear only one correct name...
Cada grupo taxonómico con sus particulares circunscripción, posición y rango podrá portar (bear) sólo un nombre correcto [aceptado]...
Código de Bacterias (Revisión 1990[30])
Principle 8. Each order or taxon of a lower rank with a given circumscription, position, and rank can bear only one correct name (...) Note 2. (i) By circumscription is meant an indication of the limits of a taxon, (ii) by position is meant the higher taxon in which a taxon is placed when there may be alternatives (see also Rule 23a), and (iii) by rank is meant level in the hierarchial sequence of taxonomic categories.
Principio 8.Cada orden o taxón de rango más bajo con unos circunscripción, posición y rango dados puede portar sólo un nombre correcto (...) Nota 2 (i) Por circunscripción se hace referencia a la indicación de los límites (limits) del taxón, (ii) por posición se hace referencia al taxón de rango más alto en el que el taxón es ubicado cuando puede haber más de una alternativa (ver también Regla 23a), y (iii) por rango se hace referencia al nivel (level) en la secuencia jerárquica de categorías taxonómicas.
Rule 23a. Each taxon above species, up to and including order, with a given circumscription, position, and rank can bear only one correct name, that is, the earliest that is in accordance with the Rules of this Code. The name of a species is a binary combination of a generic name and specific epithet (see Rule 12a). In a given position, a species can bear only one correct epithet, that is, the earliest that is in accordance with the Rules of this Code. Example: The species Haemophilus pertussis bears this name in the genus Haemophilus. If placed in the genus Bordetella, it bears the name Bordetella pertussis.
Regla 23a. Cada taxón por arriba de especie hasta orden, con sus circunscripción, posición y rango dados, pueden llevar sólo un nombre correcto, esto es, el más antiguo de los que están de acuerdo con las reglas de este Código. El nombre de una especie es una combinación binaria de un nombre genérico y un epíteto específico (ver regla 12a). En una posición dada, una especie puede llevar sólo un epíteto correcto, esto es, el más antiguo de los que están de acuerdo con las reglas de este Código. Ejemplo: la especie Haemophilus pertussis lleva este nombre en el género Haemophilus. Si se la ubica en el género Bordetella, lleva el nombre Bordetella pertussis.
Recommendation 30b. Before publication of the name and description of a new species, the examination and description should conform at least to the minimal standards (if available) required for the relevant taxon of bacteria. Note. Lists of minimal standards are being prepared for each group of bacteria by experts at the request of the Judicial Commission for consideration by the Judicial Commission and the ICSB for publication in the IJSB (see Appendix 6). Such standards include tests for the establishment of generic identity and for the diagnosis of the species, i.e., an indication of characters which would distinguish the species from others.
Recomendación 30b. Antes de la publicación del nombre y descripción de una especie nueva, la examinación y descripción deben ser conformes al menos a los estándares mínimos (si disponibles) requeridos para el taxón relevante de bacterias. Nota. La lista de estándares mínimos está siendo preparada para cada grupo de bacterias por expertos a pedido de la Comisión Judicial para consideración por la misma Comisión Judicial y el ICSB para publicación en el IJSB (ver apéndice 6). Esos estándares incluyen tests para el establecimiento de la identidad del género y para el diagnóstico de la especie, es decir, una indicación de los caracteres con los cuales distinguir la especie de las demás.
General Consideration 7. Nomenclature deals with the following: (...) Names applied to individual taxa. A taxonomic group is referred to throughout this Code as a taxon, plural taxa. "Taxonomic group" is used in this Code to refer to any group of organisms treated as a named group in a formal taxonomy; it may or may not correspond to a category.)
Consideración General 7: La nomenclatura trata con lo siguiente: (...) Nombres aplicados a taxones individuales. Un grupo taxonómico es referido a lo largo de este Código como un taxón, plural taxones (taxa). "Grupo taxonómico" es utilizado en este Código para referir cualquier grupo de organismos tratado como un grupo nombrado en una taxonomía formal, puede corresponderse o no con una categoría.
El Código de Zoología (2000[31]) utiliza como sinónimo del botánico/bacterias/unificado "circunscripción" el término aristoteliano[cita 11] "definición" (que engloba una diagnosis), en el glosario:
Código de Zoología (2000[31])
Glossary. definition, n. A statement in words that purports to give those characters which, in combination, uniquely distinguish a taxon [Arts. 12, 13]. description, n. A statement in words of taxonomic characters of a specimen or a taxon [Arts. 12, 13]. diagnosis, n. A statement in words that purports to give those characters which differentiate the taxon from other taxa with which it is likely to be confused. differentiate, v. To distinguish something (e.g. a taxon) from others [Art. 13]. See also 'definition'. Art. 12. Names published before 1931. 12.1. Requirements. To be available, every new name published before 1931 must satisfy the provisions of Article 11 and must be accompanied by a description or a definition of the taxon that it denotes, or by an indication. Art. 13. Names published after 1930. 13.1. Requirements. To be available, every new name published after 1930 must satisfy the provisions of Article 11 and must 13.1.1. be accompanied by a description or definition that states in words characters that are purported to differentiate the taxon, or 13.1.2. be accompanied by a bibliographic reference to such a published statement, even if the statement is contained in a work published before 1758, or in one that is not consistently binominal, or in one that has been suppressed by the Commission (unless the Commission has ruled that the work is to be treated as not having been published [Art. 8.7]), or 13.1.3. be proposed expressly as a new replacement name (nomen novum) for an available name, whether required by any provision of the Code or not.
Glosario. Definición. n. Una declaración (statement) en palabras que pretende dar aquellos caracteres que, en combinación, distinguen como único al taxón [Arts. 12, 13]. Descripción. n. Una declaración en palabras de los caracteres taxonómicos de un espécimen o de un taxón [Arts. 12, 13]. Diagnóstico. n. Una declaración en palabras que pretender dar aquellos caracteres que diferencien el taxón de los demás taxones con los que es probable que sea confundido. Diferenciar. v. Distinguir algo (por ejemplo un taxón) de los demás [Art. 13]. Véase también 'definición'. Art. 12. Nombres publicados antes de 1931. 12.1. Requerimientos. Para estar disponible [establecido], cada nuevo nombre publicado antes de 1931 debe satisfacer las provisiones del Art. 11 y debe estar acompañado por una descripción o una definición del taxón que denota, o por una indicación. Art. 13. Nombres publicados antes de 1930. 13.1. Requerimientos. Para estar disponible [establecido], cada nuevo nombre publicado después de 1930 debe satisfacer las provisiones del Art. 11 y debe 13.1.1. estar acompañado por una descripción o una definición que declare en palabras los caracteres que pretenden diferenciar el taxón, o 13.1.2. estar acompañado por una referencia bibliográfica a esa declaración publicada, aun si la declaración está contenida en un trabajo publicado antes de 1758, o en uno que no es consistentemente binominal, o en uno que ha sido suprimido por la Comisión (a menos que la Comisión haya reglamentado que ese trabajo sea tratado como no publicado [Art. 8.7]), o 13.1.3. ser propuesto expresamente como un nuevo nombre de reemplazo (replacement name) (nomen novum) para un nombre disponible [establecido], haya sido esto requerido por alguna provisión del Código o no.
Glossary. rank, n. The level, for nomenclatural purposes, of a taxon in a taxonomic hierarchy.
Glosario. rango. n. El nivel (level), para los propósitos nomenclaturales, de un taxón en una jerarquía taxonómica.
Glossary. taxon, (pl. taxa), n. A taxonomic unit, whether named or not: i.e. a population, or group of populations of organisms which are usually inferred to be phylogenetically related and which have characters in common which differentiate (q.v.) the unit (e.g. a geographic population, a genus, a family, an order) from other such units. A taxon encompasses all included taxa of lower rank (q.v.) and individual organisms.
Glosario. taxón, (pl. taxones), n. Una unidad taxonómica, nombrada o no: por ejemplo una población, o un grupo de poblaciones de organismos de los que usualmente se infiere que están relacionados filogenéticamente y que tienen caracteres en común que diferencia (q.v.) la unidad (por ejemplo una población geográfica, un género, una familia, un orden) de otras unidades de ese tipo. Un taxón abarca todos los taxones incluidos de rango más bajo (lower rank) (q.v.) y organismos individuales.
Biocode (2011[33])
Principle I. The BioCode governs the formation and choice of scientific names of taxa but not the circumscription, position, or rank of the taxa themselves. (...)
Principio I. El BioCode gobierna la formación y elección de los nombres científicos de los taxones pero no la circunscripción, posición, ni el rango de los mismos taxones. (...)
Principle V. Each taxon in the family group, genus group or species group with a particular circumscription, position, and rank has only one accepted name (...).
Principio V. Cada taxón en el grupo de categorías familia, en el grupo de categorías género, o en el grupo de categorías especie con unas particulares circunscripción, posición y rango tiene sólo un nombre aceptado...
↑ Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadascircunscripción posición rango
↑ El término "concepto de taxón" o "concepto taxonómico" son utilizados claramente con esa definición en:
...I would only amend the proposal of Wägele et al. to note that the taxonomic concept of the original describer is often not the concept in current use. Therefore, logic would dictate that citations should be demanded for both the paper of the original description as well as that circumscribing that actual taxonomic concept being used.
...Yo sólo modificaría la propuesta de Wägele et al. de forma de tener en cuenta que el concepto taxonómico (taxonomic concept) utilizado en la descripción original muchas veces no es el utilizado en la actualidad. Entonces, la lógica dictaría que debería demandarse que se citaran ambas publicaciones, la de la descipción original como la que circunscribió el concepto taxonómico que se está utilizando.
Johnson (2011[34])
BioNames includes two taxonomic classifications, sourced from GBIF (http://uat.gbif.org/dataset/d7dddbf4-2cf0-4f39-9b2a-bb099caae36c) and NCBI (ftp://ftp.ncbi.nih.gov/pub/taxonomy), respectively. [...] These classifications also provide an explicit definition of the scope of a taxon (i.e., the “taxon concept”).
En BioNames se incluyen dos clasificaciones, la utilizada en GBIF (http://uat.gbif.org/dataset/d7dddbf4-2cf0-4f39-9b2a-bb099caae36c) y la utilizada en NCBI (ftp://ftp.ncbi.nih.gov/pub/taxonomy). [...] Estas clasificaciones también proveen una definición explícita de los alcances del taxón (es decir, el "concepto de taxón", taxon concept).
Page (2013[35])
Ver también Franz et al. (2008[22]), On the Use of Taxonomic Concepts in Support of Biodiversity Research and Taxonomy. Taxonomic concept: The meaning of a name as specified in a particular source. Propone especificarlos con el conector sensu seguido de la fuente taxonómica.
↑ ab Existe una lucha moral (sociológica[3]) y conceptual acerca de la definición de "monofilia". Citas.
Cavalier-Smith (2010[40]):
“
-Haeckel (1868[58]) introdujo cladus como una categoría taxonómica justo debajo de subfilo, pero Huxley (1957[43], Cain y Huxley 1959[59]) dio a "clado" el significado general de cualquier rama monofilética del árbol evolutivo. Huxley lo hizo cuando contrastaba los dos procesos filogenéticos fundamentales ‘verticales’: la división cladística de los linajes y el cambio progresivo a lo largo de un linaje (de los procesos filogenéticos ‘horizontales’, sólo el sexo fue entonces apreciado, la simbiogénesis y la transferencia de genes no habían sido probadas). Huxley, como Darwin y Haeckel, correctamente enfatizaba la importancia igualitaria de la división y el cambio progresivo para entender la filogenia y la historia evolutiva. Extrañamente, la escuela de ‘sistemática filogenética’ fundada por Hennig (1966[37]) groseramente quitó importancia filogenética al cambio progresivo en comparación con la división de linajes, esto último visto por ellos como tan importantísimo que muchos devotos Hennigianos dogmáticamente insisten que los grupos ancestrales como Bacteria, Protozoa y Reptilia fueran prohibidos. Hennig llamó a los grupos basales con un origen monofilético ‘parafiéticos’ y redefinió monofilia para excluirlos e incluir sólo clados, por lo tanto redefiniéndola como incluyendo todos los descendientes de su último antecesor común. Esta redefinición de ‘clado’ es universalmente aceptada, pero la extremadamente confusa y poco sabia (unwise) redefinición de monofilia no lo es. Si bien aceptada por muchos, probablemente la mayoría (especialmente los más vociferantes y con más autoconfianza, y aquellos que prefieren no molestar a réferis anónimos, los cuales me he encontrado por docenas insistiendo equivocadamente sin argumentos razonados que los taxones parafiléticos no son permisibles en ningún caso), es firmemente rechazada por los sistemáticos evolucionistas quienes consideran que la distinción clásica entre polifilia y parafilia es mucho más importante que la distinción entre dos formas de monofilia (parafilia y holofilia, si utilizamos la terminología precisa de Ashlock (1971[60]), donde holofilético iguala a monofilético sensu Hennig).
”
Ashlock (1971[60]):
“
-Las definiciones del término monofilia en su mayoría son ambiguas porque no delimitan el ancestro común lo suficiente. Simpson (1961[15]) no la define, más bien intenta definir un "taxón nombrable". La definición de Hennig (1966[37]), que requiere que un grupo contenga todos los descendientes de un ancestro común, es precisa y útil, pero más estricta que el clásico concepto de monofilia y es aquí renombrada como holofilia. Una forma de limitar el ancestro común es requerir que sea un miembro del grupo del cual es el tallo (stem), pero esto parece requerir una apariencia macroevolutiva de los grupos. Una definición precisa de monofilia puede ser formulada si el ancestro común necesita ser un miembro sólo de forma cladística. Si un grupo puede ser mostrado como el grupo de una o más innovaciones evolutivas únicas, puede inferirse que el ancestro común más reciente también tendrá esas innovaciones, y es, entonces, cladísticamente un miembro del grupo. El ancestro necesita no haber llegado al grado de, o fenéticamente ser, un miembro del grupo. Holofilia, con el término contrastante Hennigiano parafilia, entonces se vuelven dos aspectos de la monofilia, mientras que polifilia describe un grupo cuyo ancestro común no es cladísticamente un miembro del grupo.
”
Mayr (1981[4]:nota 19):
“
-Yo utilizo el término monofilético en su sentido tradicional, como un adjetivo que califica a un taxón [a una circunscripción "evolucionista"]. Varias definiciones de monofilético han sido propuestas pero todas ellas para el mismo concepto, una cualidad que concierne a un taxón. Un taxón es monofilético si todos sus miembros son derivados del ancestro común más reciente (Haeckel 1868[58]). Los cladistas han hecho un intento de volver la situación patas arriba ubicando todos los descendientes de un ancestro en un taxón. Monofilético entonces se vuelve un adjetivo calificador de la descendencia, y un taxón no es reconocido por sus características sino por su descendencia. La transferencia de un término tan bien establecido como monofilético, a un concepto enteramente diferente es tan acientífico (unscientific) e inaceptable como si alguien tratara de "redefinir" masa, energía o gravedad asociando estos términos a conceptos enteramente nuevos. PD Ashlock (1971[60]) propuso el término holofilético para el asemblaje de descendientes de un ancestro común. Ver también PD Ashlock (1979[61] p.443).
”
↑ abc La predominancia del cladismo sobre el evolucionismo no está medida formalmente, pero se puede encontrar en la literatura que es captada como impresión por los taxónomos (aunque es evidente en los libros de texto de cursos inferiores). Cladistas como Judd et al. 2007[18]:35 sólo dicen
“
-"No todos los taxónomos utilizan una aproximación filogenética [cladista], si bien es la práctica de la mayoría".
”
Judd et al. 2007[18]:35
La minoría es la escuela evolucionista, como en Brummitt (2002[86]):
“
[Hablando de la necesidad de taxones parafiléticos] -"...De hecho, si esta audiencia es como todas las previas audiencias de las que estuve enfrente, un tercio pensará que lo que digo es tan obvio que apenas si necesita ser dicho, un tercio pensará que mi punto de vista está tan fuera de lo establecido por la convención que no necesitan molestarse en escuchar toda esta perorata en absoluto, y un tercio estará indeciso acerca de por quién tomarán partido".
”
Brummitt (2002[86])
Puede haber sesgos de apreciación debido a lo dogmática de lo que está acusada la escuela cladista, con prácticas como las mencionadas en Hörandl y Stuessy (2010[13]):
“
-"[…] muchos investigadores reaccionaron tan negativamente hacia cualquier intento de incluir grupos parafiléticos en las clasificaciones que han empezado a elevarse conflictos."
”
Hörandl y Stuessy (2010[13])
y en Cavalier-Smith (2010[40]):
“
-"[La clasificación cladista es] aceptada por muchos, lamentablemente probablemente la mayoría (especialmente … por aquéllos sin deseos de fastidiar a réferis anónimos, a quienes he encontrado por docenas insistiendo … con que los grupos parafiléticos son inadmisibles)".
”
Cavalier-Smith (2010[40])
↑ Así lo definen biólogos sistemáticos cladistas: El libro de texto "best seller" y premiado Judd et al. (edición 2007[18]) Página 5. "A phylogenetic approach demands that each taxon be a monophyletic group, defined as a group composed of an ancestor and all of its descendants:" Página 32. "The biological diversity on Earth is the result of genealogical descent with modification, and monophyletic groups owe their existence to this proccess. It is appropriate, therefore, to use monophyletic groups in biological classifications so that we may most accurately reflect this genealogical history." En la misma página: "A phylogenetic classification reflects evolutionary history and attempts to give names only to groups that are monophyletic -that is, composed of an ancestor and all its descendants-." Página 547: "Keep in mind that taxa, particularly above the level of species, represent monophyletic groups and are, as such, considered to be the products of evolution. The ranks of the taxonomic hierarchy, on the other hand, are human constructs, having relative but not absolute meaning".
↑ ab PHA Sneath (1995[67]), sobre la recepción de su filosofía en los '60: "Ciertamente muchos sistemáticos (con la excepción de los microbiólogos, quienes tenían pocos preconceptos acerca de la evolución de las bacterias) me dieron la impresión de que porque nuestras propuestas no presuponían juzgamientos filogenéticos, debían ser antievolutivas. Pocas veces comprendieron que estas propuestas podían llevar a técnicas con las cuales explorar activamente la filogenia" [...] "George Gaylord Simpson, a quien tuve el privilegio de visitar, sorpresivamente fue muy receptivo. Mientras hubiera un lugar para la filogenética, no tenía objeciones hacia estos nuevos conceptos como un primer paso. Pensó que éramos demasiado optimistas al pensar que la fenética podía traer resultados de valor en determinar inicialmente las líneas generales de la filogenia, pero no denegó ni la posibilidad ni la lógica. Simpson tenía una mente pentrativa y claramente apreció la distinción entre las relaciones fenéticas y las cladísticas." [...] "Los taxónomos de bacterias más distinguidos de la época tenían variadas visiones. En esa época había insuficiente cantidad de datos para cuestiones filogenéticas, así que el debate concernía conceptos fenéticos". [...] "Samuel Cowan [...] nos dio un gran apoyo provisto que el resultado final era una mejor identificación de bacterias". En inglés en el original: "Certainly many systematists (with the exception of microbiologists, who had few preconceptions about bacterial evolution) gave me the impression that because our proposals did not presuppose phylogenetic judgements, they must be antievolutionary. They seldom understood that these proposals could lead to techniques by which one could actively explore phylogeny." [...] "George Gaylord Simpson, whom I had the privilege of visiting [...], was surprisingly receptive. As long as there was a place for phylogenetics, he had no objection to the new concepts as a first step. He thought we were too optimistic in thinking that phenetics would yield results of value in initially determining the outlines of phylogeny, but he did not deny either the possibility or the logic. Simpson had a penetrating mind and clearly appreciated the distinction between phenetic and cladistic relationships." [...] "The most distinguished bacterial taxonomists of the time had varied views. At that time there were insufficient data for phylogenetic endeavors, so the debate concerned phenetic concepts." [...] "Samuel Cowan [...] was very supportive provided the end result was better bacterial identification."
↑ Sokal (1986[2]): "...it has been shown repeatedly that techniques developed for phenetic taxonomy, such as UPGMA clustering of correlation or distance matrices, can result in accurate estimates of the phylogeny of a group of organisms, and thus they serve as useful techniques for inferring cladistic relationships. Does this then make these techniques cladistic? Conversely, some techniques developed for phylogenetic inference have been characterized as being essentially phenetic by various investigators. In a few cases, techniques have been designated as phenetic or cladistic based solely on the philosophical orientation of their originators."
↑ ab La escuela recibió el nombre de escuela evolucionista como diferenciación del incipiente cladismo o "escuela filogenética" en la década de 1960, dos nombres que pueden prestar a confusiones. Citas.
Alessandro Minelli (1993[62]): "During the 1940s and 1950s, systematics polarized around a few leading figures, especially Ernst Mayr and George Gaylord Simpson. Later, with the development of different approaches to biological systematics, it become fashionable to identify a School of Evolutionary Systematics corresponding to the views of Mayr, Simpson and others."
Ernst Mayr (1981[4]): "Dissatisfaction with such arbitrariness and seeming absence of any carefully thought out methodology, led in the 1950's and 1960's to the establishment of two new schools of taxonomy, numerical phenetics and cladistics, and to a more explicit articulation of Darwin's methodology, now referred to as evolutionary classification."
↑ ab La clasificación en categorías altas de Cavalier-Smith fue presentada en su forma completa y actualizada por última vez en el año 1998, «A revised six-kingdom system of life», momento desde el cual fue modificada con subsiguientes publicaciones en revistas científicas por el mismo autor.
↑ PHA Sneath (1995[67]): "One cannot reconstruct phylogeny from synapomorphies if one must first know the phylogeny to recognize correctly the synapomorphies. This is logically the same as trying to select the discriminatory characters before knowing which groups are to be discriminated. One must not assume the answer in advance; to do this is not science. Phenetics may have to assume the general homologies in advance, but it does not assume the groups and then choose or alter the data so as to fit these groups. There was a strong temptation in early Hennigian studies to justify the techniques with this kind of circular argument. Hull (1984[69]) perceptively noted that the problem of paraphyletic groups is not easily disposed of by Hennigian cladistics. Platnick (1979[9]) asserted that the reason taxa seem polythetic is that the traits used are being misidentified as synapomorphies. This is an echo of an earlier argument by Remane (1956[70], see Sokal and Sneath 1963[47]:99, Sneath and Sokal 1973[68]). Yet Hull noted that this approach is close to censoring unpalatable findings. And to claim that groups cannot be polythetic is hard to credit for molecular data where amino acids and nucleotids can scarcely be misidentified. These problems soon became obvious (Colless, 1969[71]). Yet if one introduces qualifications that synapomorphies are only provisional hypotheses, recodes data to remove homoplasy, or appeals to parsimony and the like, the technique becomes iterative and certainly no longer simple. Many of these points were well discussed by Jensen (1983[72]). The concepts of Hennig were greatly expanded by his followers (Hull 1984[69], 1988[73]:244-251). But the major expositions (Eldredge and Cracraft 1980[74], Nelson and Platnick 1981[5], Wiley 1981[75]) do not in my view cover the problems adequately. The simple three-taxon method of Nelson and Platnick, the "rule of D", is not workable (Sneath 1982[76]). Recent attempts to revive the three-taxon method were heavily criticized by Harvey (1992[77]). - Noone answered my challenge to produce a simple Hennigian technique for data where relationships between organisms were not known or implied (Sneath 1982[76]:211 (first table)). If there were a simple technique there would be no arguments about phylogenies or synapomorphies, whereas such disputes continue as before. There are virtually no Hennigian analyses in microbiology or molecular taxonomy, where there are few preconceptions of the relationships of the organisms. The one attempt by Hennigian methods to determine a problem of major significance in microbiology, the relationships of blue-green algae to bacteria and eukaryotes by Humphries and Richardson (1980[78]), was merely a reiteration of traditional views based on a few allegedly important photosynthetic pigments (Sneath 1988[79], 1989[80]). By this analysis blue-green algae were most closely related to green plants, not bacteria. It gave no hint of the dramatic findings from molecular data by Woese (1981[81], 1987[82]), which indicated that blue-green algae, together with chloroplasts and mitochondria, belong to the eubacterial clade. Rogstad (1991[83]), in reviewing a collection of essays in molecular evolution (Selander et al. 1991[84]), bewailed the absence of any cladograms (synapomorphograms) in the book.".
↑ Wheeler (2004[24]): "Phylogenies remained highly speculative until Hennig (1966[37]) distinguished..."
↑ ab Así lo explicaban Camin y Sokal en 1965[50]):325: "El desarrollo de una técnica para deducir relaciones cladísticas entre los organismos parece proveer una base para una clasificación alternativa al sistema fenético expuesto por la taxonomía numérica. Como el método aquí expuesto parece ser el "filético" profesado por los taxónomos ortodoxos, puede parecer que las clasificaciones deberían ser establecidas en base a él o a un método similar. Sin embargo, ya hemos apuntado (Sokal y Camin 1965[85]) que las relaciones filéticas son siempre una composición de relaciones fenéticas, cladísticas y cronísticas no siempre claramente separadas en las mentes o escritos de los sistemáticos y que la sistemática como conjunto debería basarse en todas estas consideraciones. El grado al cual la fenética y la cladística coinciden no es conocido todavía, si bien asumimos que debe ser considerable. Como no hay un sistema operativo en este momento para combinar las relaciones fenéticas y cladísticas debemos elegir entre una base fenética o una cladística para la clasificación. Distinguimos entre "sistemática" y "clasificación", la primera incluye no sólo el estudio del orden de las cosas vivas sino también las causas y procesos que lo traen a colación, mientras que la clasificación es simplemente el arreglo de la naturaleza organizada en categorías por conveniencia de los biólogos. Por una variedad de razones, detalladas en otro lado (Sokal y Camin 1965[85]), parece ser que una base fenética es preferible para la clasificación en este sentido estricto hasta que un sistema operacional, que combine cladística y fenética pueda ser establecido".
↑ Hörandl y Stuessy (2010[13]): Some authors (e.g., Adl & al., 2005, Richter & al., 2009; Chase & Reveal, 2009) have argued for a parallel, “informal” naming of paraphyletic groups for practical use and connection to tradition, while reserving formal classification for holophyletic groups only. We reject this approach, because evolutionary classification provides a broader theoretical basis and higher information content for relationships among taxa, and therefore is more appropriate for formal classification.
↑ abc "Identificación o predicción". Mayr 1965[39]: "(Computer taxonomists) They differ from each other in basic philosophy, major objectives, and emphasis, for instance, whether the construction of a classification should be a single or multiple step procedure, how much a posteriori weighting of characters should be permitted, whether the basic objective of classification is identification or prediction, and so forth." Acerca del best-seller del mismo Mayr, Principles of Systematic Zoology, review de Richards (1970): "The fourth chapter gives a short but very satisfactory account of the history of zoological classification, a somewhat more complex subject than one might at first suppose. A gradual liberation from a priori theories was finally accomplished by Darwin, who showed that the best classifications are always those that are founded on degrees of kinship or the extent of shared gene pool. There is, however, still possibility for disagreement, since since the classification tends to serve two rather disparate purposes -the identification of specimens and the arrangement of taxa in a logical evolutionary scheme. It is desirable to keep these two aims as separate as possible, but if only for economy of space this cannot always be done in publications. Identification is usually facilitated by dichotomous keys, and the main virtue of these is that they can lead one quickly and easily to the right answer. A classification, on the other hand, may use many features that are difficult to detect in preserved material and may on the basis of characters that are weighted after study of the whole group associate forms that are superficially very different and that would not fall very near one another in a practical key."
↑ "El nombre de un organismo es la llave hacia su literatura". Frase de Davis y Heywood (1963[88]) como citada en el libro de texto de Judd et al. 2007[18]). Es una frase hecha entre taxónomos, por ejemplo también la dice Mayr (1981[4]).
↑ abc Sokal (1986[2]): "In phenetics a natural classification is defined as one in which them members of each taxon at each level are on the average more similar to each other than they are to members of other taxa at corresponding levels. (...) For the purposes of biological taxonomy, we should be concerned mainly with the formulations of the problem by Gilmour[89][90][91][92], who stated that "a system of classification is the more natural the more propositions there are that can be made regarding its constituent classes". (...) The recognition that most taxa in nature (not only biological taxa) were polythetic, led to the adoption of Gilmour naturalness as a criterion for natural classification in phenetics. Since different classifications will vary in the number of shared character states, they will consequently be more or less natural. This makes naturalness a matter of degree rather than an absolute criterion of a classification".
↑ abc Sokal (1986[2]): "In monothetic classification, classes are based on one or more invariant character states. This is the classical view of classification -now rejected. The alternative view is to regard these propositios as probabilistic, that is, class A possesses state i of character j with a probability of Pij < 1. Thus, in a polythetic taxon, membership is based on the greatest number of shared character states; no single state is either essential or sufficient to make an organism a member of the group. The recognition that most taxa in nature (not only biological taxa) were polythetic, led to the adoption of Gilmour naturalness as a criterion for natural classification in phenetics. Since different classifications will vary in the number of shared character states, they will consequently be more or less natural. This makes naturalness a matter of degree rather than an absolute criterion of a classification".
↑ Cavalier-Smith (1998[94]), traducción: "(Después de un ejemplo) ...las especies parafiléticas son por lo tanto tan naturales como las holofiléticas." - ME LO HACKEARON Y NO TENGO MÁS ACCESO AL ORIGINAL - Caso especial de las "especies biológicas" aparte, los taxones parafiléticos y holofiléticos son ambos en parte construcciones artificiales de los taxónomos (tanto si son especies asexuales como si son taxones de rango más alto). Desde esa perspectiva un taxón holofilético no es creado por el origen e una nueva sinapomorfía, sino por la decisión del sistemático de subdividir un árbol filogenético históricamente continuo en ese punto. Como comenté previamente (Cavalier-Smith 1993a[95]), cada corte de una rama holofilética necesariamente en forma simultánea genera un tallo parafilético. Si corto un árbol en piezas, cada pieza, sea ésta una pieza terminal con un corte o una pieza del tallo con dos o más, es tanto artifical como natural. Cada segmento continuo debe su continuidad a los procesos naturales, mientras que su separación de los demás es el resultado de la intervención humana, es decir artificial. Si yo agrupo piezas que había cortado que nunca estuvieron juntas en el árbol, esta unión sería totalmente no-natural (unnatural), que es la razón por la que los grupos polifiléticos se han aceptado durante tanto tiempo como mucho más artificiales que los grupos monofiléticos. Para los grupos monofiléticos (sean éstos parafiléticos como holofiléticos), su coherencia es el resultado de la continuidad genética histórica entre todos los miembros del grupo: es su discontinuidad de los demás grupos lo que es artificial (excepto por las especies biológicas). La afirmación de que un cladograma o un fenograma pueden ser convertidos en una clasificación sin cortes artificiales del continuum filogenético (Nelson & Platnick 1981[5]) es a lo sumo una media verdad espuria; los cladogramas y los fenogramas pueden ser vistos como algo tan discontinuo como las clasificaciones sólo porque los cortes en el continuum filético ya han sido hechos por taxónomos clásicos cuando crearon los taxones discretos que forman los materiales "crudos" para los análisis cladísticos y fenéticos. La afirmación implica un elemento de autoengaño miope que permite a algunos cladistas afirmar falsamente que la sistemática Hennigiana es más natural que la taxonomía clásica filética (Stuessy 1990[96]). Ninguna clasificación puede ser totalmente natural."
↑ Mayr (1981[4]:515), traducción: "No tengo duda de que los taxónomos moderados serán capaces de desarrollar una metodología ecléctica, una que contenga un balance apropiado de fenética y cladística que produzca clasificaciones mucho más "naturales" que cualquier aproximación de una-única-escuela que confíe exclusivamente en un único criterio, sea éste la similitud general (overall similarity), la parsimonia de los patrones de ramificación, o ninguna de las dos". Cita la nota 16, traducción: "Ha habido argumentos desde antes de los tiempos de Linneo sobre cómo determinar si un sistema, una clasificación, es "natural". William Whewell, en una época anterior a Darwin, ha formulado su teoría de descendencia común, expresada en el entonces prevalente consenso pragmático, "La máxima por la cual todos los sistemas que profesan ser naturales deben ser testeados es ésta: que el arreglo obtenido a partir de un set de caracteres coincida con el arreglo obtenido por otro set." (W Whewell 1840[97]) Interesantemente, la covariancia de caracteres es todavía quizás la mejor manera práctica de probar las bondades de una clasificación".
↑ Platnick (2001[10]): "...But consider for a moment what classification is all about. Is stability the primary goal of classification? Of course not! The primary goal of classification is and always has been maximal predictive power. Cladists have long argued that phylogenetic classifications are both the best summaries of the limited character information already available, and the best basis for making predictions about the much larger universe of as yet unstudied characters. Taxonomists are in the business of providing highly predictive classifications on the basis on extremely small amounts of data, and our track record of success, over the last two and a half centuries, is none too shabby".
↑ Hörandl y Stuessy (2010[13]): " For a final taxonomic decision, four main criteria should be considered in the following order: theoretical foundation based on natural processes, predictivity, information content, and practicability (e.g., Mishler, 2009). Above the species level, retention of paraphyly and holophyly is essential for the theoretical foundation of taxa through natural processes (common ancestry) and hence for predictivity. (...) The evolutionary process results in many different types of natural taxa. Groups of organisms arise from evolutionary processes. Evolution involves various kinds of processes, however, which result in different kinds of groups. By briefly reviewing these processes, we emphasize that paraphyly is a natural stage in evolution, as is holophyly, and both exist at all levels of diversification."
↑ Judd et al. (2007[18]) p.33: "¿Cómo deciden los sistemáticos qué grupos monofiléticos nombrar? No hay un set de reglas hecho Código, pero muchos criterios han sido sugeridos, y algunos son comunes a pesar de no estar del todo articulados" Wiley et al. (1991[98]), cladistas, primer capítulo, traducción: "Los métodos de clasificación de la sistemática tradicional emplean la intuición. En términos prácticos, la intuición se emplea en el peso que se le da a cada carácter." Cavalier-Smith (2010[40]), evolucionista, traducción: "El propósito de la clasificación no es "representar la genealogía" (ése es el propósito de la filogenia), sino establecer grupos nombrados de forma coherente que son sensiblemente distinguibles de otros grupos, idealmente por caracteres comunes compartidos desde sus ancestros, en un sistema jerárquico evolutivamente sólido (Cavalier-Smith 1998[94]). La taxonomía necesariamente implica una simplificación y juzgamiento de cuáles serán los caracteres enfatizados para la comprehensión humana de la biodiversidad, sin apabullar nuestros cerebros con su inmensidad. No pueden ser hechos por algoritmos delegados a las computadoras o por estudiantes graduados inexperimentados."
↑ ab Mallet (2013[120]), traducción. "Cohesión: La suma total de fuerzas o sistemas que mantienen a una especie junta. El término es usado especialmente en los conceptos de especie reproductiva (interbreeding species concept) y de especie cohesiva (cohesion species concept). Los mecanismos de cohesión incluyen mecanismos de aislamiento reproductivo en especies sexuales así como la selección estabilizadora ecológica, que puede causar cohesión aún en linajes asexuales." (En el texto) "...En una vena similar, el concepto de especie cohesiva de Templeton (1998[121]) combinó un número de ideas que competían sobre las especies. Aceptó las críticas al concepto de especie reproductiva (interbreeding), y buscó corregirla agregándole ideas de los conceptos de especie evolutiva, ecológica, de reconocimiento, y genealógica. Templeton argumentó que una combinación de cohesión ecológica y reproductiva es importante para mantener la unidad evolutiva de la especie y su integridad. Así como lo aplicó a taxones asexuales (demasiado poco sexo), Templeton también aplicó esta idea a especies como robles que hibridan frecuentemente por lo que los genes fluyen constantemente (demasiado sexo como para el concepto de especie reproductiva). Además argumentó que la separación de las genealogías es otra característica importante de las especies. Aquí nos encontramos quizás llegando al apogeo del debate sobre la especie, con esta combinación de conceptos."
Otros taxónomos, principalmente los más afines a la corriente cladista, pueden tener el mismo discurso, con otras citas y otra terminología, por ejemplo
Yeates et al. (2011[122]), traducción: "La última década ha visto verdaderos progresos en nuestra forma de pensar qué es una especie (Wiens 2007[123]). La distinción entre los conceptos de especie y el criterio o evidencia usados para delimitarlos (Hey 2006[124], de Queiroz 2007[125], Knowles y Carstens 2007[126]) han revelado que todos los conceptos de especie en el fondo son una unidad, que es que las especies son linajes (Mayden 1997[127], de Queiroz 1998[128], 2007[125]). En este sentido, muchos de los conceptos de especie en operación hoy en día representan el uso de diferente evidencia y criterios para distinguir linajes-especie (species lineages). Es instructivo que estos conceptos de especie pueden guiarnos hacia el reconocimiento de diferentes límites entre especies (Wiens y Penkrot 2002,[129] Laamanen et al. 2003,[130] Agapow et al. 2004[131]), y que muy pocos taxónomos informan el tipo de criterio o evidencia que es crítico para su concepto de especie (Schlick-Steiner et al. 2010[132]). Las dos incógnitas que representan descubrir los límites y las relaciones entre especies, están biológicamente ligadas, pero raramente están metodológicamente acopladas; sin embargo, O'Meara (2010[133]) recientemente introdujo métodos heurísticos que en conjunto estiman los límites de las especies y sus relaciones a partir de un único árbol de genes (from individual gene tree input). Estos métodos pueden mejorar nuestra comprensión de las incógnitas que contestan, pero demuestran ampliamente la complejidad del tema en teoría como en la práctica."
La cohesión —asociada a la continuidad o sobrevivencia del linaje— debe ser redefinida si se desea emplear en taxones de categoría más alta:
Nixon y Carpenter (2000[106]): "Aside from the logical conundrums of applying the concept of individuality to subclades of life, other attribues of individuality that have been expounded by its proponents are difficult or impossible to apply to clades. These include "cohesion" and "continuity", which are easily visualized in individual organisms and can be seen in species if one really squints (the acrobatic explanations of how species are "cohesive" and have continuity or are "connected" even when they occur as disjunct populations make amusing reading; cf. Mishler 1985[134], de Queiroz and Donoghue 1988[135], Baum 1998[136]). However, even the most gullible reader would have difficulty accepting the idea that aphids and blue whales show continuity and cohesion relative to their ultimate relationship in some broad clade emanating from their common ancestral worm. Argumentation along these lines merely amounts to reefinition of terms, hence obfuscation".
Por "cohesión" y "continuidad" en el tiempo como características que definen los linajes, debido a que su pool genético está integrado pero cambia a lo largo del tiempo —por lo que no necesariamente representan similitud fenotípica ni incluso genotípica— las citas tradicionales son, a partir del que se considera el iniciador del concepto de especie como linaje: Simpson 1961[15], Mayr 1963[110], Ghiselin 1974[111], Hull 1976[112], 1978[113], Wiley 1978[114], Hennig 1966[37], 1975[115], Platnick 1977[116]:97, como citados en Hull (1979[3]), un estudioso de las 3 escuelas.
↑ Biological Species Concept, cuyo principal referente es Ernst Mayr. La publicación más citada probablemente sea E Mayr (1963[110]).
↑ Phylogenetic Species Concept, los autores lo suelen citar como una forma del "concepto de especie evolutiva" de GG Simpson (1961[15]) y desarrollada en un análisis de filogenia por W Hennig y posteriormente EO Wiley (por ejemplo así lo menciona Alessandro Minelli). Publicaciones citadas son Wiley (1978[114]), Hennig (1966[37], 1975[115]). Varios autores modernos, como K de Queiroz (1998[128], 2007[125]), etc. lo proponen.
↑ En el Código de Zoología, introducción, principio 4[144]; "Nomenclatural rules are tools that are designed to provide the maximum stability compatible with taxonomic freedom.". En el Código de Botánica no se encontró que mencionaran "estabilidad".[145] En el Código de Bacterias en el primer principio del capítulo 2[146]:"The essential points in nomenclature are as follows. Aim at stability of names...". En el BioCode no se encontró "estabilidad"[147].
↑ Los principios en el Código de Zoología se encuentran en la introducción.[144] En el Código de Botánica en la División I.[145] En el Código de Bacterias en el capítulo 2.[146] En el BioCode en la División I.[147]
↑ Código de Zoología (2000[31]): Glosario. nominal taxon. A concept of a taxon which is denoted by an available name (e.g. Mollusca, Diptera, Bovidae, Papilio, Homo sapiens). Each nominal taxon in the family, genus or species groups is based on a name-bearing type (although in the latter two groups such a type may not have been actually fixed).
Código de Botánica (2012[29]): No menciona al "nominal taxon".
Código de Bacterias (Rev. 1990[30]): No menciona al "nominal taxon".
BioCode (2011[33]). 14.1. The application of names of taxa of the rank of superfamily or below, and of those names of taxa in the higher ranks that are ultimately based on generic names, is determined by means of types (name-bearing types). The unit formed by the name and its type is referred to as the nominal taxon. 14.2. A type is that element to which the name of a taxon is permanently attached, whether it be an accepted name or not.
↑ Código de Zoología. Tipificación.[148] BioCode. Tipificación.[149]
↑ ICZN faq: What are type specimens for?[31]
↑ ab Section 4. Nomenclatural Types and Their Designation. General. Rule 15. A taxon consists of one or more elements. For each named taxon of the various taxonomic categories (listed below), there shall be designated a nomenclatural type. The nomenclatural type, referred to in this Code as "type," is that element of the taxon with which the name is permanently associated. The nomenclatural type is not necessarily the most typical or representative element of the taxon. The types are dealt with in Rules 16–22. Types of the various taxonomic categories can be summarized as follows:
Table 2. Taxonomic Categories
Taxonomic category
Type
Subspecies Species}
Designated strain; in special cases the place of the type strain may be taken by a description, preserved specimen, or an illustration
Subgenus Genus}
Designated species
Subtribe Tribe
Subfamily Family
Suborder Order}
Genus on whose name the name of the higher taxon is based
Subclass Class}
One of the contained orders.
Recommendation 30a. Before publication of the name of a new species, a culture of the type strain (or, if the species is noncultivable, type material, a photograph, or an illustration) should be deposited in at least one of the permanently established culture collections from which it would be readily available. The designation allotted to the strain by the culture collection should be quoted in the published description.
↑ Código de Zoología (2000[31]). 73.3. Hapantotypes. A hapantotype (see Glossary) consisting of one or more preparations or cultures may be designated when a nominal species-group taxon of extant protistans is established. This hapantotype is the holotype of the nominal taxon. 73.3.1. A hapantotype, although consisting of a number of separate organisms, is deemed to be indivisible and cannot be restricted by lectotype selection; but 73.3.2. if a hapantotype is found to contain individuals of more than one species-group taxon, components may, by appropriate citation, be excluded from it until it contains individuals of only one species-group taxon (for the treatment of holotypes found to consist of components derived from more than one individual, see Article 73.1.5.).
↑ ab Código de Botánica (2012[29]). 8.4. Type specimens of names of taxa must be preserved permanently and may not be living organisms or cultures. However, cultures of algae and fungi, if preserved in a metabolically inactive state (e.g. by lyophilization or deep-freezing to remain alive in that inactive state), are acceptable as types. Recommendation 8B.1. Whenever practicable a living culture should be prepared from the holotype material of the name of a newly described taxon of algae or fungi and deposited in at least two institutional culture or genetic resource collections. (Such action does not obviate the requirement for a holotype specimen under Art. 8.4.)
↑ BioCode (2011[33]), Preamble. 2. The provisions of this Code shall apply to names of all kinds of organisms, whether eukaryotic or prokaryotic, fossil or non-fossil, and of fossil traces of organism (ichnotaxa), that are established (i.e., validly published or made available) and shall govern the choice when names compete among themselves or with earlier names. They shall also, and without limitation of date, provide for the establishment of co-ordinate names within rank groups, for the protection of names, as well as for their correct form. 3. Established names of organisms that are not yet covered by Adopted Lists of Protected Names are in all other respects (including their subsequent typification) governed by the International Code of Nomenclature of Bacteria (here: “bacteriological Code”), the International Code of Botanical Nomenclature (“botanical Code”) or the International Code of Zoological Nomenclature (“zoological Code”) hereafter jointly called the “Special Codes”, depending on the accepted taxonomic position of their type. 4.1. Established names are those that comply with the requirements of Art. 7-11 (see also Art. 33.2) or that, prior to the starting date defined in those Articles, were validly published or became available under the relevant Special Code. 4.2. In this Code, unless otherwise indicated, the word “name” means an established name, whether it be acceptable or unacceptable (see Art. 19). 6.1. The establishment of any family-group name is deemed to automatically establish co-ordinate names at the other rank of the family group. The co-ordinate names are formed from the same generic name and have the same authorship and date. 6.2. All subfamily names established under the zoological Code are, under the BioCode, treated as simultaneously established names of profamilies (Art. 3.2; see also Art. 24 Note 1). Note 1. Names of subfamilies established under the botanical or bacteriological Code maintain the former subfamily termination -oideae (Art. 25.1) and are intermediate in rank between profamily and tribe (Art. 3.2). 6.3. The establishment of any genus-group name is deemed to automatically establish an identical co-ordinate name at the other rank of the genus group. The co-ordinate names have the same type, authorship and date. 6.4. All names of subgenera established under the zoological Code are, under the BioCode, treated as simultaneously established names of progenera (Art. 3.2; see also Art. 26 Note 1). Note 2. Names of subgenera (subgen.) have the same form as those of other infrageneric taxa, for instance, sections (Art. 27), and are intermediate in rank between progenus and section (Art. 3.2). 6.5. The establishment of any species-group name is deemed to automatically establish a co-ordinate name at the other rank of the species group. The co-ordinate names have identical final epithets, the same type, authorship and date. 6.6. All names of subspecies established under the zoological Code are, under the BioCode, treated as simultaneously established names of prospecies (Art. 3.2; see also Art. 28 Note 1). Note 3. Names of subspecies (subsp.) have the same form as those of other infraspecific taxa, for instance, varieties (Art. 29), and are intermediate in rank between prospecies and variety (Art. 3.2).
↑ 'Código de Botánica (2012[29]). Glosario. name. A name that has been validly published, whether it is legitimate or illegitimate (Art. 6.3) (see also designation).
BioCode (2011[33]) 4.2. In this Code, unless otherwise indicated, the word “name” means an established name, whether it be acceptable or unacceptable (see Art. 19).
↑
Código de Zoología (2000[31]):
Código de Botánica (2012[29]): Glossary. heterotypic synonym (taxonomic synonym). A name based on a type different from that of another name referring to the same taxon (Art. 14.4); termed a “subjective synonym” in the International Code of Zoological Nomenclature and the International Code of Nomenclature of Bacteria (Bacteriological Code) (Art. 14.4 footnote). 14.4. A conserved name of a family or genus is conserved against all other names in the same rank based on the same type (homotypic, i.e. nomenclatural, synonyms, which are to be rejected) whether or not these are cited in the corresponding list as rejected names, and against those names based on different types (heterotypic, i.e. taxonomic, synonyms) that are listed as rejected1. A conserved name of a species is conserved against all names listed as rejected, and against all combinations based on the rejected names.
Código de Bacterias (Rev.1990[30]): Rule 24a. Note 2. These Approved Lists may contain more than one name attached to the same type (objective synonyms) since the names on the list represent those names which are considered reasonable in the present state of bacteriological nomenclature and taxonomy and represent the views of many bacteriologists who may hold different taxonomic opinions. Note 3. Synonyms may be objective synonyms (i.e., more than one name has been associated with the same type) or subjective synonyms (i.e., different names have been associated with different types that in the opinion of the bacteriologist concerned belong to the same taxon). The synonym first published is known as the senior synonym, and later synonyms are known as junior synonyms. Publication of objective synonyms in the Approved Lists does not affect bacterial nomenclature any more than does the valid publication of objective synonyms in different works in the bacteriological literature at present. Examples: Objective synonyms- Nocardia rhodochrous and Mycobacterium rhodochrous. Subjective synonyms- Leudemann (IJSB [1971] 21:240–247) regards Micromonospora fusca Jensen 1932 as a subjective synonym of Micromonospora purpureochromogenes (Waksman and Curtis 1916) Leudemann 1971. These two species have different types. Rule 41a When a species is transferred to another genus without any change of rank, the specific epithet must be retained, or if it has not been retained (in a previous publication), it must be reestablished, unless: -The resulting binary combination would be a junior homonym. Example: Pridham (1970) proposed Streptomyces aurioscleroticus for Chainia aurea Thirumalachar et al. 1966 on transfer to Streptomyces because in that genus the name Streptomyces aureus already existed. -There is available an earlier validly published and legitimate specific or subspecific epithet.
Example: Not yet found.
BioCode (2011[33]):
↑ Código de Zoología (2000[31]). Preamble. The objects of the Code are to promote stability and universality in the scientific names of animals and to ensure that the name of each taxon is unique and distinct. All its provisions and recommendations are subservient to those ends and none restricts the freedom of taxonomic thought or actions. // Priority of publication is a basic principle of zoological nomenclature; however, under conditions prescribed in the Code its application may be modified to conserve a long-accepted name in its accustomed meaning. When stability of nomenclature is threatened in an individual case, the strict application of the Code may under specified conditions be suspended by the International Commission on Zoological Nomenclature.
Código de Botánica (2012[29]). Preamble. 1. (...) This Code aims at the provision of a stable method of naming taxonomic groups, avoiding and rejecting the use of names that may cause error or ambiguity or throw science into confusion. Next in importance is the avoidance of the useless creation of names. Other considerations, such as absolute grammatical correctness, regularity or euphony of names, more or less prevailing custom, regard for persons, etc., notwithstanding their undeniable importance, are relatively accessory. DIVISION III. 4. The voting on nomenclature proposals is of two kinds: (a) a preliminary guiding mail vote and (b) a final and binding vote at the Nomenclature Section of the International Botanical Congress.
Código de Bacterias (Rev.1990[30]). General Consideration 6. 4. Provisions for emendations of Rules, for special exceptions to Rules, and for interpretation of the Rules in doubtful cases have been made by the establishment of the International Committee on Systematic Bacteriology (ICSB) and its Judicial Commission, which acts on behalf of the ICSB (see Rule 1b and Statutes of the International Committee on Systematic Bacteriology, pp. 137–158 of this volume). Opinions issued by the Judicial Commission become effective after receipt of ten or more favorable votes from Commissioners, but may be rescinded by the ICSB as provided in ICSB Statute 8c (2). The official journal of the ICSB is the International Journal of Systematic Bacteriology (IJSB), formerly the International Bulletin of Bacteriological Nomenclature and Taxonomy (IBBNT). (Some other journal could be specified by the ICSB if required. Such possible future specification is implicit in the use of "International Journal of Systematic Bacteriology" or "IJSB" throughout this Code, but is not always repeated at each mention.)
↑ abcd Código de Botánica (2012[29]): 3.1. The principal ranks of taxa in descending sequence are: kingdom (regnum), division or phylum (divisio or phylum), class (classis), order (ordo), family (familia), genus (genus), and species (species). Thus, each species is assignable to a genus, each genus to a family, etc. 4.1. The secondary ranks of taxa in descending sequence are tribe (tribus) between family and genus, section (sectio) and series (series) between genus and species, and variety (varietas) and form (forma) below species.4.2. If a greater number of ranks of taxa is desired, the terms for these are made by adding the prefix “sub-” to the terms denoting the principal or secondary ranks. An organism may thus be assigned to taxa of the following ranks (in descending sequence): kingdom (regnum), subkingdom (subregnum), division or phylum (divisio or phylum), subdivision or subphylum (subdivisio or subphylum), class (classis), subclass (subclassis), order (ordo), suborder (subordo), family (familia), subfamily (subfamilia), tribe (tribus), subtribe (subtribus), genus (genus), subgenus (subgenus), section (sectio), subsection (subsectio), series (series), subseries (subseries), species (species), subspecies (subspecies), variety (varietas), subvariety (subvarietas), form (forma), and subform (subforma). Note 1. Ranks formed by adding “sub-” to the principal ranks (Art. 3.1) may be formed and used whether or not any secondary ranks (Art. 4.1) are adopted. 4.3. Further ranks may also be intercalated or added, provided that confusion or error is not thereby introduced. Art. 16 Note 1. The terms “divisio” and “phylum”, and their equivalents in modern languages, are treated as referring to one and the same rank. When “divisio” and “phylum” are used simultaneously to denote different non-consecutive ranks, this is to be treated as informal usage of rank-denoting terms (see Art. 37 Note 1 and 37.8).
Código de Bacterias (Revisión 1990[30]) Rule 5b. The taxonomic categories above and including species which are covered by these Rules are given below in ascending taxonomic rank. Those in the left-hand column should be recognized where pertinent; those in the right-hand column are optional. The Latin equivalents are given in parentheses.
Species (Species)
. . Subgenus (Subgenus)
Genus (Genus)
. . Subtribe (Subtribus)
. . Tribe (Tribus)
. . Subfamily (Subfamilia)
Family (Familia)
. . Suborder (Subordo)
Order (Ordo)
. . Subclass (Subclassis)
Class (Classis)
Rule 5c. A species may be divided into subspecies, which are dealt with by the Rules of this Code. Variety is a synonym of subspecies; its use is not encouraged as it leads to confusion, and after publication of this Code the use of the term variety for new names will have no standing in nomenclature. Rule 5d. Taxa below the rank of subspecies (infrasubspecific subdivisions) are not covered by the Rules of this Code, but see Appendix 10.
Código de Zoología (2000[31]): No se encontraron comentarios al respecto.
BioCode (2011[33]) 2.1. The primary ranks of taxa in descending sequence are: kingdom, phylum, class, order, family, genus, and species. 3.1. Secondary ranks of taxa, when required, include in descending sequence: domain above kingdom, tribe between family and genus, section and series between genus and species, and variety and form below species. 3.2. Should an even greater number of ranks of taxa be desired, the terms for these are made by adding the prefixes super-, pro-, or sub- (sub- being below pro-) to non-prefixed terms denoting principal or secondary ranks. 11.1. Only if the corresponding genus or species name is established can the name of a subordinate taxon be established (but see Art. 30.2).
↑ "The establishment of the Bulletin of Zoological Nomenclature in 1942 meant that zoologists had a single place in which to present applications to resolve problems that could not satisfactorily be resolved under provision of the code. The ICZN subsequently publishes and votes on these applications. In 1953, the International Association of Plant Taxonomy was established and with it a similar venue for publication of proposals to modify the code, the journal Taxon (although today the journal is devoted to much more than just nomenclatural issues).". Knapp et al. 2004[156]
↑ Revista Taxon 62(1), febrero 2013, "Guidelines for authors": "The convention of only italicizing names at the rank of genus and below is followed, except for nomenclature papers and proposals to conserve or reject names in which all scientific names are italicized, regardless of rank. http://www.iapt-taxon.org/downloads/guidelines_authors.pdf[153]
↑ ab Código de Zoología: "Los autores deberían ejercitar un razonable cuidado y consideración en la formación de nombres nuevos para asegurarse de que son elegidos con sus subsecuentes usuarios en mente y que, mientras sea posible, sean apropiados, compactos, eufónicos, fáciles de memorizar y no causen ofensa".[157]
↑ abcdefghijklmnñop Código de Botánica (2012[29]). DIVISIÓN II. Reglas y Recomendaciones. Capítulo III. Nomenclatura de los taxones de acuerdo con su rango. Sección 1. Taxones por arriba del rango de familia. 16.1. The name of a taxon above the rank of family is treated as a noun in the plural and is written with an initial capital letter. Such names may be either (a) automatically typified names (Art. 10.7), formed from the name of an included genus in the same way as family names (Art. 18.1; but see Art. 16.4) by adding the appropriate rank-denoting termination (Art. 16.3 and 17.1), preceded by the connecting vowel –o– if the termination begins with a consonant; or (b) descriptive names, not so formed, which may be used unchanged at different ranks. Ex.1. Automatically typified names above the rank of family: Lycopodiophyta, based on Lycopodium; Magnoliophyta, based on Magnolia; Gnetophytina, based on Gnetum; Pinopsida, based on Pinus; Marattiidae, based on Marattia; Caryophyllidae and Caryophyllales, based on Caryophyllus; Fucales, based on Fucus; Bromeliineae, based on Bromelia. Ex.2. Descriptive names above the rank of family: Anthophyta, Chlorophyta, Lycophyta, Parietales; Ascomycota, Ascomycotina, Ascomycetes; Angiospermae, Centrospermae, Coniferae, Enantioblastae, Gymnospermae. (...) Note 2. The principle of priority does not apply above the rank of family (Art. 11.10; but see Rec. 16A). Recommendation 16A 16A.1. In choosing among typified names for a taxon above the rank of family, authors should generally follow the principle of priority.
Código de Bacterias (Revisión 1990[30]) Section 3. Naming of Taxa. Rule 8. The name of each taxon above the rank of order is a Latin or latinized word preferably in conformity with Recommendation 6. It is based by choice on a combination of characters of the taxon or from a single character of outstanding importance. Example: Kingdom-Procaryotae; Class-Schizomycetes. Rule 9. The name of a taxon between subclass and genus is formed by the addition of the appropriate suffix to the stem of the name of the type genus (see Rule 15). These suffixes are as follows: Table 1. Suffixes for Categories. Rank, Suffix, Example. Order, -ales, Pseudomonadales. Suborder, -ineae, Pseudomonadineae. Family, -aceae, Pseudomonadaceae. Subfamily, -oideae, Pseudomonadoideae. Tribe, -eae, Pseudomonadeae. Subtribe, -inae, Pseudomonadinae
Section 4. Nomenclatural Types and Their Designation. General. Rule 15. A taxon consists of one or more elements. For each named taxon of the various taxonomic categories (listed below), there shall be designated a nomenclatural type. The nomenclatural type, referred to in this Code as "type," is that element of the taxon with which the name is permanently associated. The nomenclatural type is not necessarily the most typical or representative element of the taxon. The types are dealt with in Rules 16–22. Types of the various taxonomic categories can be summarized as follows:
Table 2. Taxonomic Categories.
{Taxonomic category: Type}
{Subspecies, Species; Designated strain; in special cases the place of the type strain may be taken by a description, preserved specimen, or an illustration.}
{Subgenus, Genus: Designated species.}
{Subtribe, Tribe, Subfamily, Family, Suborder, Order: Genus on whose name the name of the higher taxon is based.}
{Subclass, Class: One of the contained orders.}
Código de Zoología (2000[31]): Article 29. Family-group names. 29.1. Formation of family-group names. A family-group name is formed by adding to the stem of the name [Art. 29.3] of the type genus, or to the entire name of the type genus [see Article 29.6], a suffix as specified in Article 29.2.
BioCode (2011[33]): Article 23. 23.1. Names of taxa above the rank of family are treated as nouns in the plural and are written with a capital initial letter. They may be either (a)typified names (see Art. 14.1) that are formed by adding a termination denoting their rank to the genitive singular stem of a generic name, or exceptionally to the whole name; or (b) typeless (“descriptive”) names that are formed differently, apply to taxa defined by circumscription, and may be used unchanged at different ranks. 23.2. For typified names, the name of a subphylum that includes the type of the accepted name of a phylum, the name of a subclass that includes the type of the accepted name of a class, or the name of a suborder that includes the type of the accepted name of an order, are to be based on the same type. 23.3. The typified name of a phylum or subphylum is formed from the same generic name as an acceptable name of an included class. The phylum name termination is -mycota for fungi, -phyta for other botanical taxa, and -zoa for animals. The subphylum name termination is -mycotinafor fungi, -phytina for other botanical taxa, and -zoina for animals. 23.4. The typified name of a class or subclass is formed from the same generic name as an acceptable name of an included order. The class name termination is -mycetes for fungi, -phyceae for algae, and -opsida for other botanical taxa and all animals. The subclass name termination is -mycetidae for fungi, -phycidae for algae, -idae for other botanical taxa, and -zoidae for animals. 23.5. The typified name of an order, suborder, or superfamily is formed from the same generic name as an acceptable name of an included family. For all groups, the order name termination is -ales, the suborder name termination -ineae, and the superfamily name termination -oidea. 23.6. The name of a taxon above the rank of family may not have the termination -virinae, -virales, or -viridae, because these terminations are reserved for the names of viral taxa (see Pre. 4). Note 1. Names of taxa above the rank of family that do not conform to the standards set out in Art. 23.3-23.6 are acceptable as descriptive names (Art. 23.1(b)). 23.7. When a typified suprafamilial name is published with a Latin termination not agreeing with the provisions of this Article, the termination is changed to accord with it, but the name retains its authorship and date. Note 2. For suprafamilial names of ambiregnal taxa (Rec. 10A, footnote), the alternative use of the terminations -mycota or -phyta and -zoa (for phyla), -mycotinaor -phytina and -zoina (for subphyla), -mycetes or -phyceae and -opsida (for classes), -mycetidae or -phycidae and -zoidae (for subclasses) is authorised, irrespective of the Special Code otherwise used by a given author. Recommendation 23A 23A.1. The terminations provided in Art. 23.3-23.5 should not be used in typeless, descriptive names of any rank above family.
↑ abcdefghijkl Código de Botánica (2012[29]) 16.3. Automatically typified names end as follows: the name of a division or phylum ends in -phyta, unless it is referable to the algae or fungi in which case it ends in -phycota or -mycota, respectively; the name of a subdivision or subphylum ends in -phytina, unless it is referable to the algae or fungi in which case it ends in -phycotina or -mycotina, respectively; the name of a class in the algae ends in -phyceae, and of a subclass in -phycidae; the name of a class in the fungi ends in -mycetes, and of a subclass in -mycetidae; the name of a class in the plants ends in -opsida, and of a subclass in -idae (but not -viridae). Automatically typified names not in accordance with these terminations or those in Art. 17.1 are to be corrected, without change of the author citation or date of publication (see Art. 32.2). However, if such names are published with a non-Latin termination they are not validly published.
↑ abcdef Hay recomendaciones en el BioCode.
Código de Zoología (2000[31]): No posee recomendaciones en relación al nombre por arriba del grupo familia.
BioCode (2011[33]): Article 23. 23.1. Names of taxa above the rank of family are treated as nouns in the plural and are written with a capital initial letter. They may be either (a) typified names (see Art. 14.1) that are formed by adding a termination denoting their rank to the genitive singular stem of a generic name, or exceptionally to the whole name; or (b) typeless (“descriptive”) names that are formed differently, apply to taxa defined by circumscription, and may be used unchanged at different ranks. 23.2. For typified names, the name of a subphylum that includes the type of the accepted name of a phylum, the name of a subclass that includes the type of the accepted name of a class, or the name of a suborder that includes the type of the accepted name of an order, are to be based on the same type. 23.3. The typified name of a phylum or subphylum is formed from the same generic name as an acceptable name of an included class. The phylum name termination is -mycota for fungi, -phyta for other botanical taxa, and -zoa for animals. The subphylum name termination is -mycotina for fungi, -phytina for other botanical taxa, and -zoina for animals. 23.4. The typified name of a class or subclass is formed from the same generic name as an acceptable name of an included order. The class name termination is -mycetes for fungi, -phyceae for algae, and -opsida for other botanical taxa and all animals. The subclass name termination is -mycetidae for fungi, -phycidae for algae, -idae for other botanical taxa, and -zoidae for animals. 23.5. The typified name of an order, suborder, or superfamily is formed from the same generic name as an acceptable name of an included family. For all groups, the order name termination is -ales, the suborder name termination -ineae, and the superfamily name termination -oidea. 23.6. The name of a taxon above the rank of family may not have the termination -virinae, -virales, or -viridae, because these terminations are reserved for the names of viral taxa (see Pre. 4). Note 1. Names of taxa above the rank of family that do not conform to the standards set out in Art. 23.3-23.6 are acceptable as descriptive names (Art. 23.1(b)). 23.7. When a typified suprafamilial name is published with a Latin termination not agreeing with the provisions of this Article, the termination is changed to accord with it, but the name retains its authorship and date. Note 2. For suprafamilial names of ambiregnal taxa (Rec. 10A, footnote), the alternative use of the terminations -mycota or -phyta and -zoa (for phyla), -mycotinaor -phytina and -zoina (for subphyla), -mycetes or -phyceae and -opsida (for classes), -mycetidae or -phycidae and -zoidae (for subclasses) is authorised, irrespective of the Special Code otherwise used by a given author. Recommendation 23A 23A.1. The terminations provided in Art. 23.3-23.5 should not be used in typeless, descriptive names of any rank above family.
↑ ab Código de Bacterias (revisión 1990[30]) Rule 8. The name of each taxon above the rank of order is a Latin or latinized word preferably in conformity with Recommendation 6. It is based by choice on a combination of characters of the taxon or from a single character of outstanding importance. Example: Kingdom-Procaryotae; Class-Schizomycetes.
↑ ab Código de Botánica (2012[29]) 17.1. Automatically typified names of orders or suborders are to end in -ales (but not -virales) and -ineae, respectively (see Art. 16.3 and 32.2). 17.2. Names intended as names of orders, but published with their rank denoted by a term such as “cohors”, “nixus”, “alliance”, or “Reihe” instead of “order”, are treated as having been published as names of orders. Recommendation 17A 17A.1. A new name should not be published for an order for which a name already exists that is based on the same type as the name of an included a family.
↑ abcdef Código de Bacterias (Revisión 1990[30]) Rule 9. The name of a taxon between subclass and genus is formed by the addition of the appropriate suffix to the stem of the name of the type genus (see Rule 15). These suffixes are as follows:
Table 1. Suffixes for Categories.
Rank Suffix Example
Order -ales Pseudomonadales
Suborder -ineae Pseudomonadineae
Family -aceae Pseudomonadaceae
Subfamily -oideae Pseudomonadoideae
Tribe -eae Pseudomonadeae
Subtribe -inae Pseudomonadinae.
↑ abcde Código de Zoología (2000[31]): Article 29. Family-group names. 29.1. Formation of family-group names. A family-group name is formed by adding to the stem of the name [Art. 29.3] of the type genus, or to the entire name of the type genus [see Article 29.6], a suffix as specified in Article 29.2. 29.2. Suffixes for family-group names. The suffix -OIDEA is used for a superfamily name, -IDAE for a family name, -INAE for a subfamily name, -INI for the name of a tribe, and -INA for the name of a subtribe. These suffixes must not be used at other family-group ranks. The suffixes of names for taxa at other ranks in the family-group are not regulated. 29.2.1. Names in the genus and species groups which have endings identical with those of the suffixes of family-group names are not affected by this Article.
↑ Código de Botánica (2012[29]) 18.1. The name of a family is a plural adjective used as a noun; it is formed from the genitive singular of a name of an included genus by replacing the genitive singular inflection (Latin -ae, -i, -us, -is; transcribed Greek -ou, -os, -es, -as, or -ous, and its equivalent -eos) with the termination -aceae (but see Art. 18.5). For generic names of non-classical origin, when analogy with classical names is insufficient to determine the genitive singular, -aceae is added to the full word. Likewise, when formation from the genitive singular of a generic name results in a homonym, -aceae may be added to the nominative singular. For generic names with alternative genitives the one implicitly used by the original author must be maintained, except that the genitive of names ending in -opsis is always -opsidis.
↑ Código de Botánica (2012[29]) 19.1. The name of a subfamily is a plural adjective used as a noun; it is formed in the same manner as the name of a family (Art. 18.1) but by adding the termination -oideae instead of -aceae.
↑ ab Código de Botánica (2012[29]) 19.3. A tribe is designated in a similar manner, with the termination -eae, and a subtribe similarly with the termination -inae (but not -virinae).
↑ Mayr 1978[208] citando a Sneath 1962: Monothetic: 1 feature neccesary and sufficient OR set of features severally neccesary and jointly sufficient for inclusion in a group.
↑ Mayr 1965[39]: "The choice of the term "numerical taxonomy" for this very special taxonomic theory (Sokal and Sneath 1963[47]) seems rather unfortunate, when we remember how far back (about 200 years) good taxonomists have insisted on using the "greatest possible number of characters". That only a consideration of the totality of characters can lead to a sound classification was already expressed by Buffon and Adanson in the middle of the 18th century in their criticism of the Linnean method. Blumenback wrote in 1779, "according to our concept of a natural method we must consider not merely a few selected but all external characteristics indeed the total habitus of animals". In 1788, Merrem expressed similar ideas when he said, "genera, families, orders and classes must be based on the similarity of all parts of the body and not on individual (single) characters". Considering how many different kinds of numerical taxonomies have existed in the past or actually exist today, some with philosophies diametrically opposed to that of Sokal and Sneath, it would seem advantageous to reer to their particular ideology by a more appropriate term. The basic tenet of this school seems to be the de-emphasis of all information other than the calculated resemblance (phenetic distance) of taxa..."
↑ "Pequeño lab" en contraposición a "big lab" sensu de Carvalho et al. (2008[230]): "...a palpable sense of isolation affecting many taxonomists nested in university departments wherein the majority of faculty are biochemists, physiologists, molecular biologists, and other ‘big lab’ professionals who usually have greater amounts of funding and frequently work (and publish) in teams...".
↑ Wheeler (2004[24]): "La taxonomía es una ciencia excitante, no un servicio de identificación para otros científicos, si bien la necesidad de identificaciones confiables de especies es una justificación mayor para apoyar esta ciencia."
↑ abc de Queiroz (2000[27]): "Contrary to Stuessy's view, taxon names are defined in both traditional and phylogenetic systems of nomenclature (de Queiroz 1997[375]). In the traditional system of nomenclature, taxon names are defined in terms of types and taxonomic categories. For example, the definition of the name Asteraceae is "the taxon containing the genus Aster that is assigned to the family category". In the phylogenetic system, taxon names are defined in terms of specifiers and their common ancestry relationships. For example, a node-based phylogenetic definition of the name Asteraceae might be "the clade stemming from the most recent common ancestor of Barnadesia and Aster."
Referencias citadas
↑ Brian S. Everitt, Sabine Landau, Morven Leese, Daniel Stahl. 2011. «Dendrograms and other tree representations.» Cluster analysis. Wiley
↑ abcdefghi Sokal, Robert R. (1986). «Phenetic Taxonomy: Theory and Methods». Annu. Rev. Ecol. Syst. (17): 423-442. Consultado el 28 de agosto de 2007.
↑ abcdefgh Hull, D. L. (1979). The limits of cladism. Systematic Biology, 28(4):416-440.
↑ abcdefghij Mayr, E. (1981). Biological classification: toward a synthesis of opposing methodologies. Science 214:510-516. http://www.joelvelasco.net/teaching/systematics/mayr%2081%20-%20biological%20classification%20-%20toward%20a%20synthesis.pdf
↑ abcdefghi Nelson, G, y y N. Platnick. 1981. Systematics and biogeography. Columbia University Press. Nueva York.
↑ ab Patterson, C. 1982. Morphological characters and homology. En: Problems of phylogenetic reconstruction. ed. KA Jorsey, AE Friday. pp. 21-74. New York: Academic
↑ Beatty, J. (1982). Classes and cladists. Systematic Biology, 31(1), 25-34.
↑ Brady RH. 1982. Theoretica issues and "pattern cladistics". Syst. Zool. 31:286-291.
↑ ab Platnick, N. 1979. Philosophy and the transformation of cladistics. Syst. Zool. 28:537-546.
↑ abcdef Platnick, N. I. (2001). From Cladograms to Classifications: The Road to DePhylocode. Oral presentation at Systematics Association AGM, Linnean Society of London, organized by the Systematics Association. http://www.systass.org/archive/events-archive/2001/platnick.pdf
↑ Ebach, M.C. & Williams, D. 2004. Classification. Taxon 53: 791–794. http://www.jstor.org/stable/4135452
↑ ab Podani, J. (2010). Monophyly and paraphyly: A discourse without end?. Taxon, 59(4), 1011-1015. http://www.ingentaconnect.com/content/iapt/tax/2010/00000059/00000004/art00002
↑ abcde Hörandl, Stuessy. 2010. «Paraphyletic groups as natural units of biological classification.» Taxon
↑ Por ejemplo Stuessy, T. F., & König, C. (2009). Classification should not be constrained solely by branching topology in a cladistic context. Taxon, 58(2), 347-348. "Consistencia lógica" en respuesta a Wiley (2009).
↑ abcdefghijk Simpson, G. G. (1961). Principles of animal taxonomy (Columbia University Press edición). Nueva York. ISBN 0-231-02427-4.
↑ Mayr, E.; Ashlock, P. D. (1991). Principles of Systematic Zoology. 2ª ed. (McGraw-Hill edición). Nueva York. ISBN 0-07-041144-1. La referencia utiliza el parámetro obsoleto|coautores=(ayuda) Decía K de Queiroz (1992): "En 1969, el libro de Mayr, Principles of Systematic Zoology fue probablemente el mejor texto de sistemática y el mejor libro de referencia disponible (Mihener 1969), y retuvo un lugar prominente entre los de su estilo por años después de su publicación. Cuando fue publicado por primera vez, estaba actualizado, era considerado una autoridad en la materia, y era el más completo, y aún más de 20 años después de su apogeo continúo usando mi copia frecuentemente. Si bien muchos libros excelentes sobre el tema han sido publicados después, sólo el de Wiley (1981) Phylogenetics, sirve a tantos propósitos diferentes. Pero un montón ha sucedido desde 1969. Ningún libro sobre sistemática publicado hace tanto tiempo es actual hoy en día. Consecuentemente, la publicación de una edición revisada de Principles of Systematic Zoology se espera que sea un nuevo estándar en este campo."
↑ ab Richards OW (1970) Book Reviews: Principles of Systematic Zoology Ernst Mayr. McGraw-Hill, New York, 1969. xiv + 434 pp., illus. $ 12.50. Science 167(3924):1477-1478. http://www.sciencemag.org/content/167/3924/1477.extract
↑ abcdefghijklmnñopqrstuvwxyzaaabacadaeafagahaiajakalamanañaoapaqarasatauavawaxayazbabbbcbdbebfbg Judd, W. S.; Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. (2007). Plant systematics: a phylogenetic approach, Third Edition (Sinauer Axxoc edition). USA. ISBN 0-87893-403-0.
↑ ab Mayr, Ernst. 1942. Systematics and the Origin of Species. p.17 "...the basic principle of the rules, that of stability and permanence,..."
↑ abcdefghi Michael G. Simpson. 2005. Plant Systematics. Elsevier Academic Press. La segunda edición es del 2010.
↑ ab QD Wheeler (2008) Introductory. Toward the New Taxonomy. En: QD Wheeler (ed. 2008). The New Taxonomy. Capítulo 1. https://books.google.com.ar/books?hl=en&id=Ykf8RALCGyUC&pg=PP1
↑ ab Franz, N. M., Peet, R. K., & Weakley, A. S. (2008). On the use of taxonomic concepts in support of biodiversity research and taxonomy. Systematics Association Special Volume, 76, 63. http://www.researchgate.net/profile/Alan_Weakley/publication/228951937_On_the_use_of_taxonomic_concepts_in_support_of_biodiversity_research_and_taxonomy/links/0deec5216ad710dbaf000000.pdf
↑ abcdefghijklmnñopqrs Landrum, L. R. (2001). What has happened to descriptive systematics? What would make it thrive?. Systematic Botany, 26(2), 438-442. https://www.researchgate.net/profile/Leslie_Landrum/publication/298134141_What_has_happened_to_descriptive_systematics_What_would_make_it_thrive/links/56f4427008ae95e8b6d065a2.pdf
↑ abcdefghijklmnñopqrst Wheeler, Q. D. 2004. Taxonomic triage and the poverty of phylogeny. Phil. Trans. R. Soc. Lond. B 359. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1693342/pdf/15253345.pdf
↑ Brummitt, R. K. & Sosef, M. S. M. 1998 Paraphyletic taxa are inherent in Linnaean classification: a reply to Freudenstein. Taxon 47, 411–412.
↑ ab Stuessy, T. F. (2000). Taxon names are not defined. Taxon, 231-233.
↑ abc de Queiroz, K. (2000). The definitions of taxon names: a reply to Stuessy. Taxon, 533-536. http://vertebrates.si.edu/herps/herps_pdfs/deQueiroz_pdfs/2000deqtaxon.pdf
↑ abcd K. de Queiroz. 2012. «Biological Nomenclature from Linnaeus to the PhyloCode.» En Bell, C. J. (ed.) The Herpetological Legacy of Linnaeus: A Celebration of the Linnaean Tercentenary, Bibliotheca Herpetologica 9(1-2):135-145. http://hydrodictyon.eeb.uconn.edu/eebedia/images/5/55/2012DeQueiroz.pdf
↑ abcdefghijklmnñopq J. MCNEILL, Chairman, F. R. BARRIE, W. R. BUCK, V. DEMOULIN, W. GREUTER, D. L. HAWKSWORTH, P. S. HERENDEEN, S. KNAPP, K. MARHOLD, J. PRADO, W. F. PRUD'HOMME VAN REINE, G. F. SMITH, J. H. WIERSEMA, Members, N. J. TURLAND, Secretary of the Editorial Committee: International Code of Botanical Nomenclature (Melbourne Code) adoptado por el Eighteenth International Botanical Congress Melbourne, Australia, julio de 2011. Publ. 2012. Regnum Vegetabile 154. A.R.G. Gantner Verlag KG. ISBN 978-3-87429-425-6 http://www.iapt-taxon.org/nomen/main.php
↑ abcdefghijklmnñ Editado por SP Lapage, PHA Sneath, EF Lessel, VBD Skerman, HPR Seeliger, and WA Clark. 1992. International Code of Nomenclature of Bacteria. Bacteriological Code, 1990 Revision. International Union of Microbiological Societies. Washington (DC): ASM Press; 1992. ISBN 1-55581-039-X http://www.ncbi.nlm.nih.gov/books/NBK8817/
↑ abcdefghijklmnñopq International Commission on Zoological Nomenclature. 2000. International Code of Zoological Nomenclature. International Trust for Zoological Nomenclature.
↑ Hull, DL. 1965. The effect of essentialism on taxonomy - Two thousand years of stasis (I). Br. J. Philos. Sci., 15:314-326.
↑ abcdefghi Greuter, W., Garrity, G., Hawksworth, D. L., Jahn, R., Kirk, P. M., Knapp, S., ... & Tindall, B. J. (2011). Draft BioCode (2011): principles and rules regulating the naming of organisms. http://www.bionomenclature.net/biocode2011.html Publicado en:
G. Garrity et al., “Draft BioCode (2011): Principles and Rules Regulating the Naming of Organisms”, Bulletin of Zoological Nomenclature, vol. 68, no. 1, pp. 10-28, 2011. http://iczn.org/content/draft-biocode-2011-principles-and-rules-regulating-naming-organisms
Greuter, W., Garrity, G., Hawksworth, D. L., Jahn, R., Kirk, P. M., Knapp, S., ... & Tindall, B. J. (2011). Draft BioCode (2011): principles and rules regulating the naming of organisms. Taxon, 201-212. http://www.biotaxa.org/Bionomina/article/download/166/264
↑ abcde Johnson Norman F. (2012) A collaborative, integrated and electronic future for taxonomy. Invertebrate Systematics 25, 471–475. http://dx.doi.org/10.1071/IS11052 http://www.publish.csiro.au/view/journals/dsp_journal_fulltext.cfm?nid=120&f=IS11052
↑ abcdefgh Page, R. D. (2013). BioNames: linking taxonomy, texts, and trees. PeerJ, 1, e190. https://peerj.com/articles/190/
↑ abc Hennig, W. (1950). Grundzuege einer Theorie der phylogenetischen Systematik. Véase la siguiente edición, escrita en inglés, de 1966.
↑ abcdefghij Hennig W. 1966. Phylogenetic Systematics. (es una traducción/ampliación de su libro en alemán de 1950, ver también resumen de conceptos, previo a la traducción, de 1965, Phylogenetic systematics. Ann. Rev. Entomol. 10:97–116. Hay una "excelente" traducción al español injustamente poco conocida según Dupuis 1984 del manuscrito de 1961 publicada en 1968: Hennig W, 1968. Elementos de una sistemática filogenética. Buenos Aires, Ed. Univ. 353 pp.
↑ abc E Mayr (1982) The Growth of Biological Thought: Diversity, Evolution and Inheritance. Cambridge, Mass: Belknap. 974 pp.
↑ abcde Mayr, Ernst. 1965. Numerical phenetics and taxonomic theory. Syst. Zool. 14:73-97. http://www.jstor.org/stable/2411730
↑ abcdefghijklmn Cavalier-Smith, T. 2010. «Deep phylogeny, ancestral groups and the four ages of life.» Phil. Tans. R. Soc B. 365, 111-132. (contenido libre bajo Creative Commons Attribution License)
↑ ab C Dupuis (1984) Willi Hennig's Impact on Taxonomic Thought. Ann. Rev. Ecol. Syst. 1984 15:1-24. http://www.annualreviews.org/doi/pdf/10.1146/annurev.es.15.110184.000245
↑ Falta la referencia con el autor Rensch, probablemente se refiere a un trabajo muy leído de Bernhard Rensch que fue traducido al inglés en una edición revisada, como Evolution above the Species Level en 1960 (Columbia University Press, New York). GG Simpson en su review del libro comenta que el original alemán kladogenese fue traducido al inglés como kladogenesis y un anglicismo correcto sería cladogenesis (GG Simpson 1961, http://www.jstor.org/stable/2405847
↑ ab JS Huxley (1957) The three types of evolutionary process. Nature 180:454-455
↑ ab JS Huxley (1958) Evolutionary processes and taxonomy with special references to grades. Uppsala Univ. Arsskr. 1958 (6):21-38.
↑ L Cuénot (1940) Remarques sur un essai d'arbre généalogique du regne animal. C. R. Acad. Sci. 210:23-27.
↑ abc AJ Cain y GA Harrison. 1960. Phyletic weighting. Proc. Zool. Soc. London 135:1-31.
↑ abcdef RR Sokal, PHA Sneath. 1963. Principles of Numerical Taxonomy. Freeman, San Francisco. 359 pp.
↑ ab W Hennig (1957) Systemati und Phylogenese. En: Bericht über die Hundertjahrfeier d. Dtsch. Entom. Gesells. Berlin, 30 Sept-5 Oct. 1956, pp. 50-71. Berlin: Akad.
↑ E Mayr (1965) Classification and phylogeny. Am. Zool. 5:165-74.
↑ ab JH Camin y RR Sokal (1965) A method for deducing branching sequences in phylogeny. Evolution 19:311-326. http://www.jstor.org/stable/2406441
↑ WW Moss (1983) Report on NATO Advanced Study Institute on Numerical Taxonomy. Syst. Zool. 32:76-83.
↑ RR Sokal (1983) A phylogenetic analysis of the Caminalcules. I-IV. Syst. Zool. 32:159-184; 185-201; 248-58; 259-75.
↑ Farris, J. S. (1979). «The Information Content of the Phylogenetic System». Syst. Zool. (28): 458-519. doi:10.2307/2412562. Consultado el 28 de agosto de 2007.
↑ Humphries, C. J. y L. R. Parenti. 1986. Cladistic biogeography. Clarendon, Oxford.
↑ Brooks, D. R. y D. A. McLennan. 1991. Phylogeny, ecology and behavior. University of Chicago Press, Chicago.
↑ Forey, P. L., C. J. Humphries, I. L. Kitching, R. W. Scotland, D. J. Siebert, y D. M. Williams. 1992. Cladistics: a practical course in systematics. Oxford University Press. Oxford.
↑ Cracraft, J. y M. J. Donoghue. 2004. Assembling the tree of life. Oxford University Press, Oxford y Nueva York.
↑ ab Haeckel E. 1868 Naturliche Schöpfungsgeschichte. Berlin, Germany: Reimer.
↑ Cain A. J. Huxley J. S. 1959 Clades and grades. En: Function and taxonomic importance (ed. Cain A. J.), pp. 21–22. London, UK: The Systematics Association.
↑ abc Ashlock P. D. 1971 Monophyly and associated terms. Syst. Zool. 20, 63–69.(doi:10.2307/2412223) http://sysbio.oxfordjournals.org/content/20/1/63?ijkey=40c030534faf0ca3b7963b18070ef0897febb3f1&keytype2=tf_ipsecsha
↑ ab Ashlock, P. D. (1979). «An Evolutionary Systematist's View of Classification». Syst. Zool. (28): 441-450. doi:10.2307/2412559. Consultado el 28 de agosto de 2007. Judd et al. (2007): Presentación de un método explícito y fácil de seguir para construir una clasificación taxonómica evolucionista.
↑ Alessandro Minelli. 1993. Biological Systematics. The state of the art. Springer Netherlands. 406 pp.
↑ Stevens, PF. 1986. Evolutionary classification in botany, 1960-1985. J. Arnold Arbor. 67:313-339.
↑ R. H. Whittaker, 1969. New Concepts of Kingdoms of Organisms. Science Vol. 163. no. 3863, pp. 150 – 160 DOI: 10.1126/science.163.3863.150 «Copia archivada». Archivado desde el original el 13 de agosto de 2012. Consultado el 8 de febrero de 2013.
↑ L Margulis. 1971. Whittaker's Five Kingdoms of Organisms: Minor revisions suggested by considerations of the origin of mitosis. Evolution 25 (1) - quizás sea un paper falso y sea correcta la cita de Curtis y Barnes Biología (Whittaker y Margulis 1978).
↑ Takhtajan, Armen. 2009. Flowering Plants, 2nd edition. Springer-Verlag
↑ abcdefg Sneath, P. H. (1995). «Thirty years of numerical taxonomy.» Systematic Biology, 44(3), 281-298.
↑ ab Sneath, P. H. A.; R. R. Sokal (1973). Numerical Taxonomy (Freeman edición). San Francisco. ISBN 0-7167-0697-0. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ ab Hull, DL. 1984. Cladistic theory: Hypotheses that blur and grow. Páginas 5-23 en Cladistics (T Duncan y TF Stuessy, eds.) Columbia Univ. Press, New York.
↑ Remane, A. 1956. Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik. Theoretische Morphologie und Systematik, 2nd edition, Volume 1. Geest und Pörtig, Leipzig.
↑ Colles, DH. 1969. The interpretation of Hennig's "Phylogenetic systematics" -A reply to Dr. Schlee. Syst. Zool. 18:134-144.
↑ Jensen, RJ. 1983. A practical view of numerical taxonomy, or should I be a pheneticist or a cladist? Páginas 53-71 in Numerical taxonomy (J Felsenstein, ed.) Springer-Verlag, Berlin.
↑ Hull, DL. 1988. Science as a process. Univ. Chicago Press, Chicago.
↑ Eldredge, N y J Cracraft. 1980. Phylogenetic patterns and the evolutionary process. Columbia Univ. Press, New York.
↑ Wiley, EO. 1981. Phylogenetics. Wiley, New York.
↑ ab Sneath, PHA. 1982. Review of Systematics and biogeography: Cladistics and vicariance, by G. Nelson and N. Platnick. Syst. Zool. 31:208-217.
↑ Harvey, AW. 1992. Three-taxon statements: More precisely, an abuse of parsimony? Cladistics 8:345-354.
↑ Humphries, CJ y PM Richardson. 1980. Hennig's method and phytochemistry. Páginas 353-378 en: Chemosystematics: Principles and practice (FA Bisby, JG Vaughan y CA Wright, eds). Academic Press, London.
↑ Sneath, PHA. 1988. The phenetic and cladistic approaches. Páginas 252-273 en Prospects in systematics (DL Hawksworth, ed.) Clarendon Press, Oxford, England.
↑ Sneath, PHA. Analysis and interpretation of sequence data for bacterial systematics: The view of a numerical taxonomist. Syst. Appl. Microbiol. 12:15-31.
↑ Woese, C. 1981. Archaebacteria. Sci. Am. 244:94-106.
↑ Woese, C. 1987. Bacterial evolution. Microbiol. Rev. 51:221-271.
↑ Rogstad, SH. 1991. Evolution at the molecular level. Cladistics 7:307-310.
↑ Selander, RK; AG Clark, y TS Whittam (eds.) 1991. Evolution at the molecular level. Sinauer, Sunderland, Massachusetts.
↑ ab RR Sokal, JH Camin (1965). The two taxonomies: areas of agreement and conflict. Systematic Zoology, 176-195. http://www.jstor.org/stable/2411548
↑ abc Brummitt. 2002. «How to chop up a tree.» Taxon
↑ ab Adl, S. M., Simpson, A. G., Farmer, M. A., Andersen, R. A., Anderson, O. R., Barta, J. R., … & Taylor, M. F. (2005). «The new higher level classification of eukaryotes with emphasis on the taxonomy of protists.» Journal of Eukaryotic Microbiology, 52(5), 399-451.
Adl, S. M., Simpson, A. G., Farmer, M. A., Andersen, R. A., Anderson, O. R., Barta, J. R., ... & James, T. Y. (2005). The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. Journal of Eukaryotic Microbiology, 52(5), 399-451. http://www.treesearch.fs.fed.us/pubs/download/30101.pdf
↑ ab Davis, P. D.; Heywood, V. H. (1963). Principles of Angiosperm Taxonomy (Van Nostrand edición). Nueva York. ISBN 0-05-000824-2. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ Gilmour, JSL. 1937. A taxonomic problem. Nature 139:1040-42.
↑ Gilmour, JSL. 1940. Taxonomy and philosophy. En: The New Systematics, ed. J. Huxley, pp. 461-74. Oxford: Clarendon. 583 pp.
↑ Gilmour, JSL. 1951. The development of taxonomic theory since 1851. Nature 168:400-2.
↑ Gilmour, JSL. 1961. Taxonomy. En: Contemporary Botanical Thought, ed. A. M. MacLeod, L. S. Cobley, pp. 27-45. Edinburgh: Oliver and Boyd. 197 pp.
↑ abc de Queiroz, K.; Gauthier, J. (1990). «Phylogeny as a Central Principle in Taxonomy: Phylogenetic Definitions of Taxon Names». Syst. Zool. (39): 307-322. doi:10.2307/2992353. Consultado el 28 de agosto de 2007. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ ab Cavalier-Smith, T. 1998. A revised six-kingdoms of life. Biol. Rev. 73: 203-266. http://aulavirtual.maristasmediterranea.com/pluginfile.php/60812/mod_resource/content/1/Teor%C3%ADa%20de%20los%20seis%20Reinos.%20Cavalier%20Smith.pdf
↑ Cavalier-Smith, T. (1993). Kingdom Protozoa and its 18 phyla. Microbiological Reviews 57, 953–994.
↑ Stuessy, T. F. (1990). Plant Taxonomy. Columbia University Press, New York. Segunda edición en: Stuessy, T. F. 2009. Plant taxonomy: The systematic evaluation of comparative data. 2nd ed. Columbia University Press, New York.
↑ W Whewell, 1840. 'Philos. Inductive Sci. 1:521.
↑ EO Wiley, D Siegel-Causey, DR Brooks, VA Funk. 1991. The compleat cladist: a primer of phylogenetic procedures. Vol. 19. Lawrence, Kansas: Museum of Natural History, University of Kansas, 1991. http://faculty.uml.edu/rhochberg/hochberglab/courses/arthropods/Wiley%20et%20al%201991.pdf
Archivado el 13 de junio de 2015 en la Wayback Machine.
↑ abcdefghi Dayrat, B. 2005. Towards Integrative Taxonomy. Biological Journal of the Linnean Society 85.
↑ abcdefgh Will, K. W., BD Mishler, QD Wheeler. 2005. The Perils of DNA Barcoding and the Need for Integrative Taxonomy. Systematic Biology 54.
↑ Vences, M., et al. 2013. «To name or not to name: Criteria to promote economy of change in Linnaean classification schemes.» Zootaxa, [S.l.], v. 3636, n. 2, pp. 201-244, apr. 2013. ISSN 1175-5334. [1]. Consultado el 10 de junio de 2015. doi: http://dx.doi.org/10.11646/zootaxa.3636.2.1.
↑ Stevens, P. F. (1997). Dransfield, J., Coode, M. J. E., y Simpson, D. A., ed. Plant diversity in Malesia III: Proceedings of the 3rd International Flora Malesiana Symposium 1995 (Royal Botanical Gardens edición). Kew, Londres. pp. 295-319. ISBN 1-900347-42-3.
↑ ab Nevo, Wasser. 2004. Evolutionary theory and processes. p. 295
↑ Wiley, EO; Lieberman (2011). Phylogenetics, Theory and Practice of Phylogenetic Systematics, Second Edition (Wiley-Blackwell edición). USA. ISBN 0-470-90596-4.
↑ Moore, G. (1998). A comparison of traditional and phylogenetic nomenclature. Taxon 47, 561–579. http://www.jstor.org/stable/1223578 Citado en relación al tema de las categorías taxonómicas por Nixon y Carpenter (2000) Cladistics.
↑ abcdefg Nixon, K. C., y J.M. Carpenter. 2000. On the Other "Phylogenetic Systematics". Cladistics 16, 298–318 (2000) doi:10.1006/clad.2000.0137 http://155.97.32.9/~mhaber/Documents/Course%20Readings/NixonCarpenter_Clad2000.pdf
Archivado el 7 de septiembre de 2015 en la Wayback Machine. Paper retomado en:
Nixon, K.C., Carpenter, J.M., Stevenson, D.W., 2003. The PhyloCode is fatally flawed, and the “Linnaean system” can easily be fixed. Bot. Rev. 69, 111-120. http://www.botany.wisc.edu/courses/botany_940/05PhyloCode/papers/NixonCarpen2003.pdf
↑ ab Godfray, HCJ, S Knapp. 2004. Introduction. Phil. Trans. R. Soc. Lond. B 359.
↑ Stevens, P. F. (1997). «How to Interpret Botanical Classifications: Suggestions from History» (pdf). Bioscience (47): 243-250. doi:10.2307/1313078. Consultado el 28 de agosto de 2007. (enlace roto disponible en Internet Archive; véase el historial y la última versión).
↑ De Queiroz, K. (2006). «The PhyloCode and the distinction between taxonomy and nomenclature.» Systematic biology, 55(1), 160-162.
↑ abcd Mayr, E. 1963. Animal Species and Evolution. Harvard University Press, Cambridge.
↑ ab M Ghiselin 1974. A radical solution to the species problem. Syst. Zool. 23:536-544.
↑ ab DL Hull 1976. Are species really individuals? Syst. Zool. 25:174-191.
↑ ab DL Hull. 1978. A matter of individuality. Phil. Sci. 45:335-360.
↑ abc EO Wiley 1978. The evolutionary species concept reconsidered. Syst. Zool. 27:17-26.
↑ abc W Hennig 1975. "Cladistic analysis or cladistic classification?": a reply to Ernst Mayr. Syst. Zool. 24:244-256.
↑ ab NI Platnick 1977. Review of concepts of species. Syst. Zool. 26:96-98.
↑ abcdefghijklmnñop Isaac, N. J., Mallet, J., & Mace, G. M. (2004). Taxonomic inflation: its influence on macroecology and conservation. Trends in Ecology & Evolution, 19(9), 464-469. http://www.ucl.ac.uk/taxome/jim/pap/isaac04.pdf
↑ Son las secciones en las que suelen estar divididos los libros de Ecología General (un sinónimo de "interacciones biológicas") como el de Krebs.
↑ P.ej. en Akçacaya et al. 1999. Applied Population Ecology. Principles and Computer Exercises, using RAMAS(R) Ecolab. Second Edition. Sinauer Associates. Inc.
↑ Mallet J. 2013. Species, concepts of. En: SA Levin, ed. Encyclopedia of Biodiversity. Volume 6. Waltham, Mass.: Academic Press, 679-691. http://www.ucl.ac.uk/taxome/jim/pap/mallet%202013%20sp%20concepts%20inEnc%20Biodiv%20edited.pdf
↑ Templeton AR (1998) Species and speciation. Geography, population structure, ecology, and gene trees. En: Howard DJ y Berlocher SH (eds.) Endless Forms. Species and Speciation. pp. 32-43. New York: Oxford University Press.
↑ abcd DK Yeates, A Seago, L Nelson, SL Cameron, L Joseph, JWH Trueman (2010) «Integrative taxonomy, or iterative taxonomy?» Systematic Entomology (2011)36:209-217. http://www.researchgate.net/publication/229957015_Integrative_taxonomy_or_iterative_taxonomy/file/e0b495189e43f722e0.pdf
↑ J Wiens (2007) Global diversity patterns: evolutionary and ecological causes. Natural history museums and institutions in the 21st Century: impact on our common future. Proceedings of the Buffon International Symposium, 18-19 October, pp.34-39. Museum National d'Histoire Naturelle, Paris.
↑ J Hey (2006) On the failure of modern species concepts. Trends in Ecology and Evolution 21:447-450.
↑ abc K De Queiroz (2007) Species concepts and species delimitation. Systematic Biology 56:879-886.
↑ LL Knowles y BC Carstens (2007) Delimiting species without monophyletic gene trees. Systematic Biology 56:887-895.
↑ RL Mayden (1997) A hierarchy of species conepts: the denouement in the saa of the species problem. Species: The Units of Biodiversity (ed. by MF Claridge, HA Dawah and MR Wilson). pp.381-424. Chapman and Hall Ltd, London.
↑ ab K De Queiroz (1998) The general lineage concept of species, species criteria, and the process of speciation: a conceptual unification and terminological recommendations. Endless Forms: Species and Speciation (ed. by DJ Howard and SH berlocher), pp. 57-75. Oxford University Press, Oxford.
↑ JJ Wiens, TA Penkrot (2002) Delimiting species using DNA and morphological variation and discordant species limits in spiny lizards (Sceloporus). Systematic Biology 51:69-91.
↑ TR Laamanen, FT Petersen y R Meier (2003) Kelp flies and species concepts -the case of Coelopa frigida (Fabricius, 1805) and C. nebularum Aldrich, 1929 (Diptera: Coelopidae). Journal of Zoological Systematics and Evolurionay Research 41:127-136.
↑ PM Agapow, ORP Bininda-Emonds, KA Crandall, JL Gittleman, GM Mace, JC Marshall y A Purvis (2004) The impact of species concept on biodiversity studies. Quarterly Review of Biology 79:161-179.
↑ ab BC Schlick-Steiner, FM Steiner, B Seifert, C Stauffer, E Christian, RH Crozier (2010) Integrative taxonomy: a multisource approach to exploring biodiversity. Annual Review of Entomology 55:421-438.
↑ BC O'Meara (2010) New heuristic methods for joint species delimitation and species tree inference. Systematic Biology 59:59-73.
↑ Mishler, B. D. (1985). The morphological, developmental and phylogenetic basis of species concepts in bryophytes. Bryologist 88,207–214
↑ de Queiroz, K., and Donoghue, M. J. (1988). Phylogenetic systematics and the species problem. Cladistics 4, 317–338
↑ Baum, D. A. (1998). Individuality and the existence of species through time. Syst. Biol. 47, 641–653.
↑ abcdefgh Boenigk J, Ereshefsky M, Hoef-Emden K, Mallet J, Bass D. 2012. Concepts in protistology: species definitions and boundaries. European Journal of Protistology 48: 96-102 http://www.ucl.ac.uk/taxome/jim/pap/boenigk%20et%20al%202012%20protist%20species.pdf This paper summarises the Symposium ‘Concepts in Protistology’, during the VI European Congress of Protistology, Berlin, 25–29 July 2011.
↑ abcdefghijklmnñop J. Mallet; Willmott, K. (2003). «Taxonomy: Renaissance or Tower of Babel?» (pdf). Trends Ecol. Evol. 18 (2): 57-59. Consultado el 28 de agosto de 2007. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ abcdefghijklmnñopqr Linnaeus, C. (1751). Philosophia Botanica (Kiesewetter edición). Estocolmo. ISBN 0-19-850122-6. , edición citada en Judd et al. (2007) Plant Systematics capítulo 3.
↑ E Mayr (1969) Principles of systematic zoology. McGraw-Hill, New York.
↑ Brickell, C.D. et al. (eds) (2009). «International Code of Nomenclature for Cultivated Plants». Scripta Horticulturae (8th edición) (International Society of Horticultural Science) 10: 1-184. ISBN 978-0-643-09440-6.
↑ International Standards for Naming Pathovars of Phytopathogenic Bacteria ("plant pathogen Standards") http://www.isppweb.org/about_tppb_naming.asp.
↑ International Code of Virus Classification and Nomenclature: http://ictvonline.org/codeOfVirusClassification_2012.asp
↑ ab Los principios en ICZN Code - Introducción. http://www.nhm.ac.uk/hosted-sites/iczn/code/index.jsp?nfv=true&booksection=introduction
↑ ab Código de Botánica. División I. Principios. http://www.iapt-taxon.org/nomen/main.php?page=div1
↑ ab Código de Bacterias. Capítulo 2. Principios. http://www.ncbi.nlm.nih.gov/books/n/icnb/A171/
↑ ab BioCode. División I. Principios. http://archive.bgbm.org/biodivinf/docs/biocode2011/biocode2011.htm#DIVISIONI
↑ ICZN Code Article 61. Principio de Tipificación. http://www.nhm.ac.uk/hosted-sites/iczn/code/index.jsp?article=61&nfv=true
↑ Section 4. Typification. http://www.bionomenclature.net/biocode2011.html
↑ ab "During 2001–2010, the total number of new animal taxa fluctuated between 18,106 and 20,709 per year according to Zoological Record (Fig. 6A)" http://www.mapress.com/zootaxa/2011/f/zt02896p007.pdf
↑ abcdefghijklmnñ David L. HAWKSWORTH (2011) BioCode 2011. Introducción. http://www.bionomenclature.net/biocode2011.html
↑ International Commission on Zoological Nomenclature. «The Bulletin of Zoological Nomenclature» (en inglés). Archivado desde el original el 23 de mayo de 2009. Consultado el 5 de julio de 2009.
↑ ab International Association for Plant Taxonomy. «Taxon» (en inglés). Archivado desde el original el 2 de octubre de 2013. Consultado el 28 de julio de 2013.
↑ International Journal of Systematic and Evolutionary Microbiology (IJSEM) ijs.sgmjournals.org/content/journal/ijsem
↑ McNeill (1997) citado en Patterson et al. (2006) Taxonomic indexing: extending the role of Taxonomy.
↑ abc Knapp, S., Lamas, G., Lughadha, E. N., y Novarino, G. (2004). Stability or stasis in the names of organisms: the evolving codes of nomenclature. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 359(1444), 611-622.
↑ Recomendación 25c del Código de Zoología, traducción. http://www.nhm.ac.uk/hosted-sites/iczn/code/index.jsp?article=29&nfv=#2
↑ «Curiosities of Biological Nomenclature». Enero de 2008. Consultado el 19 de mayo de 2008. La referencia utiliza el parámetro obsoleto|mes=(ayuda)
↑ ab Berendsohn, W. G. (1995). The concept of "potential taxa" in databases. Taxon, 207-212. http://www.bgbm.fu-berlin.de/BioDivInf/Docs/PotentialTaxa/PotentialTaxa.pdf
↑ abc Page, RDM. 2008. Biodiversity informatics: the challenge of linking data and the role of shared identifiers. Brief Bioinform (2008) 9 (5): 345-354 first published online April 29, 2008 doi:10.1093/bib/bbn022 http://bib.oxfordjournals.org/content/9/5/345.full
↑ Vane-Wright, R.I. (2003). «Indifferent Philosophy versus Almighty Authority: on Consistency, Consensus and Unitary Taxonomy». Syst. Biodiv. doi:10.1017/S1477200003001063. Archivado desde el original el 14 de diciembre de 2007. Consultado el 28 de agosto de 2007.
↑ Will et al. (2005, The Perils of DNA Barcoding...), las descripciones de especies nunca están terminadas, porque siempre se están agregando caracteres y poblaciones nuevos que obligan a retestearlas, y a ver cómo evolucionan en sus caracteres y en su distribución en el tiempo. Ejemplos de estos cambios se encuentran documentados en las colecciones más antiguas en museos y jardines botánicos.
↑ NSF Systematics and Biodiversity Science Cluster (SBS). 1. Biodiversity: Discovery & Analysis program. http://www.nsf.gov/funding/pgm_summ.jsp?pims_id=503666&org=DEB&from=home Accedido el 4 de septiembre del 2016. En: NSF National Science Foundation. Where Discoveries Begin. http://www.nsf.gov
↑ como en el caso comentado por Soltis et al. 2005 sobre la ubicación filogenética de Petermanniaceae
↑ por ejemplo en: Wiley EO, Lieberman. 2011. "Historical Biogeogaphy." En: Phylogenetics. Theory and Practice of Phylogenetic Systematics. Second Edition. Wiley-Blackwell. Capítulo 9.
↑ Leadlay, Jury. 2006. Taxonomy And Plant Conservation: The Cornerstone of the Conservation and the Sustainable Use of Plants. p. 297.
↑ Resultados de la búsqueda de plantas tóxicas para el ganado. En: "Validación de una estrategia de autogestión para la prevención y control de enfermedades en el ganado de pequeños productores agropecuarios". Médicas Veterinarias Paula Lucía Olaizola y Eloisa Luján Ferro. Sesión Pública Extraordinaria del 28 de octubre de 2011. Entrega del Premio: “Biogénesis Bagó - versión 2011”. Anales de la Acad. Nac. de Agron. y Vet. Tomo LXV (2011) p. 342/550
↑ abc Burrows, Tyrl. 2012. Toxic Plants of North America. Second Edition. Wiley. p. 1280.
↑ Berman, 2012. Taxonomic Guide to Infectious Diseases: Understanding the Biologic Classes of Pathogenic Organisms. Elsevier Academic Press.
↑ ab Gibb, Oseto. 2010. Arthropod collection and identification: Laboratory and field techniques.
↑ como en Masuh, Héctor. 2008. Re-emergence of dengue in Argentina: Historical development and future challenges
Archivado el 2 de octubre de 2013 en la Wayback Machine..
↑ Página 12. Entevista con la entomóloga Adriana Oliva: el caso Carrasco. http://www.pagina12.com.ar/diario/suplementos/futuro/subnotas/1036-140-2004-12-18.html
↑ Entomología forense en la Argentina (con entrevista a la Dra Adriana Oliva)
Archivado el 11 de septiembre de 2012 en la Wayback Machine.. 2006. Revista del Hospital Ramos Mejía.
↑ GBIF. Data about Argentina. http://www.gbif.org/country/AR/summary Accedido el 07-09-2016.
↑ BOLD. http://v3.boldsystems.org BOLD Accedido el 07-09-2016.
↑ abcde Bortolus, A. (2008). Error cascades in the biological sciences: the unwanted consequences of using bad taxonomy in ecology. AMBIO: A Journal of the Human Environment 37(2), 114-118.
↑ Vink, C. J., Paquin, P., & Cruickshank, R. H. (2012). Taxonomy and irreproducible biological science. BioScience, 62(5), 451-452.
↑ ab Ruedas, L. A., Salazar-Bravo, J., Dragoo, J. W., & Yates, T. L. (2000). The importance of being earnest: what, if anything, constitutes a “specimen examined?”. Molecular phylogenetics and evolution, 17(1), 129-132.
↑ ««Animal Research Review Panel and NSW Department of Primary Industries - Animal Welfare Branch.» "Collection of voucher specimens." En: ANIMAL ETHICS INFOLINK. ARRP policies and guidelines. Wildlife research.».
↑ ab IZCN Code. Appendix B. General Reccomendations. "In the case of new species-group names, when the name-bearing types consist of one or more preserved specimens their deposition in a collection cited by name and location is mandatory [Art. 16.4]; wherever possible this collection should be housed in an appropriate institution." http://www.nhm.ac.uk/hosted-sites/iczn/code/index.jsp
↑ De Salle, Giribet, Wheeler. 2002. "Documentation and Voucher Specimens". En: Techniques in Molecular Systematics and Evolution. Springer
↑ Wiley EO, Lieberman. 2011. "Specimens and Curation." En: Phylogenetics. Theory and Practice of Phylogenetic Systematics. Second Edition. Wiley-Blackwell. Capítulo 10.
↑ abc Hawthorne y Lawrence. 2012. Plant Identification: Creating User-Friendly Field Guides for Biodiversity Management. Capítulo 5: «Identification: Keys and other access methods.»
↑ Judd et al. (2007) p.54, en su glosario de morfología de plantas: "Muchos, si no todos, los términos esquematizados aquí deberían ser considerados meramente puntos convenientes a lo largo de un continuum de variación en la forma. Si bien son útiles para la comunicación, condiciones intermedias serán encontradas". En inglés en el original: "Many, if not all, of the terms outlined here should be considered merely convenient points along a continuum of variation in form. Although they are useful in communication, intermediate conditions will be encountered".
Bajo el nombre "Caracteres tipológicos". Por ejemplo en Rato et al. (2016:Rato, C., Harris, D. J., Carranza, S., Machado, L., & Perera, A. (2016). The taxonomy of the Tarentola mauritanica species complex (Gekkota: Phyllodactylidae): Bayesian species delimitation supports six candidate species. Molecular phylogenetics and evolution, 94, 271-278.): "...la especies han sido identificadas y descriptas utilizando caracteres morfológicos, tanto tipológicos como cuantitativos..." En inglés en el original: "...species have been identified and described using morphological traits, either typological or quantitative..."
↑ Askevold, I. S. y C. W. O'Brien. 1994. DELTA, an invaluable computer program for generation of taxonomic monographs. Ann. Entomol. Soc. Am. 87: 1-16.
↑ Dallwitz, M. J. 1993. DELTA and INTKEY. En Advances in computer methods for systematic biology: artificial intelligence, databases, computer vision. R. Fortuner (ed.), 287-296. The Johns Hopkins University Press, Baltimore, MD.
↑ Dallwitz, M. J. 2000 onward. A comparison of interactive identification programs. http://delta-intkey.com/www/comparison.htm
↑ Dallwitz, M. J., T. A. Paine, y E. J. Zurcher. 2002 en adelante. Principles of Interactive Keys. http://delta-intkey.com/www/interactivekeys.htm
↑ Pankhurst, R. J. 1978. Biological Identification. University Park Press. Baltimore, MD.
↑ Pankhurst, R. J. 1991. Practical Taxonomic Computing. Cambridge University Press. Cambridge.
↑ Watson. L. y M. J. Dallwitz. 1991. The families of angiosperms: Automated descriptions, with interactive identification and information retrieval. Aust. Syst. Bot. 4: 681-695
↑ Watson, L. y M. J. Dallwitz. 1993. The families of flowering plants: Interactive identifications and information retrieval (CD-ROM version 1.0 for MS-DOS). CSIRO, Melbourne, Australia.
↑ Norton GA. DJ Patterson. M Schneider. 2012. LucID: A multimedia educational tool for identification and diagnostics. IJISME. http://ojs-prod.library.usyd.edu.au/index.php/CAL/article/view/6141
↑ Edwards, M. y D. R. Morse. 1995. The potential for computer-aided identification in biodiversity research. Trends Ecol. Evol. 10: 152-158.
↑ «http://www.colby.edu/info.tech/B1211/PlantFamilyD.html». (enlace roto disponible en Internet Archive; véase el historial y la última versión).
↑ «http://www.discoverlife.org».
↑ antweb http://www.antweb.org/index.jsp
↑ guía de campo de saltamontes comunes del oeste http://www.sdvc.uwyo.edu/grasshopper/fieldgde.htm
↑ mariposas y polillas de norteamérica http://www.butterfliesandmoths.org
↑ Farr, D. F. 2006. On-line keys: More than just paper on the web. Taxon 55: 589-596.
↑ National Institute of Invasive Species Science Thomas J. Stohlgren Science Program Director, Natural Resources Ecology Lab Colorado State University. 2006. Measuring Plant Diversity: Lessons from the field. Oxford University Press.
↑ abcde CHC Lyal, AL Weitzman 2004. Taxonomy: Exploring the Expediment. Science.
↑ Hey, J. (2001). «The mind of the species problem». Trends in Ecology and Evolution 16 (7): 326-329.|fechaacceso=requiere|url=(ayuda)
↑ Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadasLinneo 1753
↑ Greene, E. L. (1983). «Landmarks of Botanical History, 2 ols.». En F. N. Egerton. Stanford University Press (Stanford, C.A.). fue escrito casi 100 años antes de su publicación, pero la mitad del libro fue publicado por primera vez recién en 1981 (cuidadosamente editado entonces), este trabajo cubre la botánica hasta el siglo XVI, si bien autores más tardíos, como Tournefort, también fueron incluidos. (Judd et al. 2007)
↑ ab Adanson, M. (1763-1764). Familles des Plantes. 2 vols.vol.1 pp xxii-xxvii (Vincent edición). París. ISBN 3-7682-0345-X. citado en Judd et al. 2007
↑ Broberg, G. 1975. Homo sapiens 156-194. Citado por Schiebinger, L. 1993. Why Mammals are called mammals: Gender Politics in Eighteen-Century Natural History. The American Historical Review 98(2)
↑ Mayr, E. (1978). Origin and history of some terms in systematic and evolutionary biology. Systematic Zoology, 27(1), 83-88. http://www.jstor.org/stable/2412818
↑ de Jussieu, A. -L. (1789). Genera Plantarum secundum ordines naturales disposita, juxta methodum in hortu regio Parisiensi exaratum, anno M. DCC. LXXIV. (Hérissant and Barrois edición). París.
↑ Candolle, Augustin Pyrame de (1813). Théorie élémentaire de la botanique, ou Exposition du principe de la classification naturelle et de l'art de décrire et d'étudier les végétaux. Paris: Déterville.
↑ ab Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadasMayr et al. 1953
↑ Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadasHuxley 1940
↑ Strickland, H.E., Henslow, J.S., Phillips, J., Shuckard, W.E., Richardson, J.B., Waterhouse, G.R., Owen, R., Yarrell, W., Jenyns, L., Darwin, C., Broderip, W.J. & Westwood, J.O. (1842). Report of a Committee appointed "to consider the rules by which the nomenclature of Zoology may be established on a uniform and permanent basis." John Murray, London, for the British Association for the Advancement of Science. [También publicado en 1843 como: Series of propositions for rendering the nomenclature of zoology uniform and permanent, being a report of a Committee for the con-sideration of the subject appointed by the British Association for the Advancement of Science. Annals and Magazin of Natural History, 11, 259–275].
↑ Mayr, E. 1942. Systematics and the Origin of Species, from the viewpoint of a zoologist. Harvard University Press, 334 pp.
↑ ab Brickell, C.D. et al. (eds) (2009). "International Code of Nomenclature for Cultivated Plants". Scripta Horticulturae (International Society of Horticultural Science) 10: 1–184. ISBN 978-0-643-09440-6.
http://www.actahort.org/chronica/pdf/sh_10.pdf
«Copia archivada». Archivado desde el original el 15 de octubre de 2005. Consultado el 3 de octubre de 2005.
↑ The Angiosperm Phylogeny Group III ("APG III", en orden alfabético: Brigitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis y Peter F. Stevens, además colaboraron Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang y Sue Zmarzty) (2009). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.» (pdf). Botanical Journal of the Linnean Society (161): 105-121.
↑ abc «PhyloCode» (en inglés). Consultado el 6 de febrero de 2013.
↑ Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadasDe Queiroz y Gauthier 1990
↑ Dall, W.H. (1877) Nomenclature of Zoology and Botany. A Report to the American Association for the Advancement of Science at the Nashville Meeting, August 31, 1877. Salem Press, Salem, 56 pp.
↑ Savory, T. 1962. Naming the living world. John Wiley and Sons, New York.
↑ Allen, D. E. 1976. The naturalist in Britain: A social history. A. Lane, Londres.
↑ Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadasJudd et al. 2002
↑ Nieto Olarte, M. (2003). «Historia Natural y la Apropiación del Nuevo Mundo en la Ilustración Española». Bull. Inst. Fr. Ètudes Andines 3 (32): 417-429.|fechaacceso=requiere|url=(ayuda) Para las frases textuales de Ulloa cita como fuente a Pagden, 1993: 132
↑ Muelas, L. (1997). «La Patente del Yagé». Semillas (9). Archivado desde el original el 21 de diciembre de 2007. Consultado el 28 de agosto de 2007.
↑ abcde Wheeeler, QD, PH Raven, y EO Wilson. 2004. Taxonomy: impediment or expedient? Science 303.
↑ abc Padial, J. M., Miralles, A., De la Riva, I., & Vences, M. (2010). Review: The integrative future of taxonomy. Front Zool, 7, 1-14. http://www.biomedcentral.com/content/pdf/1742-9994-7-16.pdf
↑ ab Steele, C., Butler, L., & Kingsley, D. (2006). The publishing imperative: the pervasive influence of publication metrics. Learned Publishing, 19(4), 277-290. http://dx.doi.org/10.1087/095315106778690751
↑ Wilson, E. O. 2000. On the future of conservation biology. Conservation Biology 14(1): 1–3. 2000. ‘‘Phylogenetic reconstruction, currently the dominating focus of systematics, obviously is worth doing, but more scientifically important and far more urgent for human welfare is the description and mapping of the world biota.’’ (E. O. Wilson, 2000) como citado en Landrum (2001).
↑ abcdefgh Godfray, H. C. J. (2002). Challenges for taxonomy. Nature, 417(6884), 17-19. http://limnoreferences.missouristate.edu/assets/limnoreferences/Godfray_2002_taxonomy_on_the_web.pdf
Archivado el 2 de julio de 2010 en la Wayback Machine.
↑ abcdefghijklm De Carvalho, M. R., Bockmann, F. A., Amorim, D. S., & Brandao, C. R. F. (2008). Systematics must embrace comparative biology and evolution, not speed and automation. Evolutionary Biology, 35(2), 150-157. http://www.researchgate.net/profile/Flavio_Bockmann/publication/225326984_Systematics_must_Embrace_Comparative_Biology_and_Evolution_not_Speed_and_Automation/links/00b4951781dc1c3c24000000.pdf
↑ Gotelli, N. J. 2004. A taxonomic wish-list for community ecology. Phil. Trans. R. Soc. Lond. B 359. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1693344/pdf/15253346.pdf
↑ Mace, G. M. 2004. The role of taxonomy in species conservation. Phil. Trans. R. Soc. Lond. B http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1693347/pdf/15253356.pdf
↑ abcdefgh Ebach, M. C., Valdecasas, A. G., & Wheeler, Q. D. (2011). Impediments to taxonomy and users of taxonomy: accessibility and impact evaluation. Cladistics, 27(5), 550-557. http://onlinelibrary.wiley.com/doi/10.1111/j.1096-0031.2011.00348.x/abstract?userIsAuthenticated=false&deniedAccessCustomisedMessage=
↑ abcde Evenhuis, N. L. (2007). Helping solve the “other” taxonomic impediment: completing the eight steps to total enlightenment and taxonomic nirvana. Zootaxa, 1407, 3-12. http://mapress.com/zootaxa/2007f/zt01407p012.pdf
↑ abcde Werner, Y. L. (2006). The case of impact factor versus taxonomy: a proposal. Journal of Natural History, 40(21-22), 1285-1286. http://www.tandfonline.com/doi/full/10.1080/00222930600903660.
↑ abcdefg Thessen A, Patterson D (2011) Data issues in the life sciences. ZooKeys 150: 15-51. doi: 10.3897/zookeys.150.1766 http://zookeys.pensoft.net/articles.php?id=3041
↑ abcdefghijklm Agnarsson I, M Kuntner. 2007. Taxonomy in a Changing World: Seeking Solutions for a Science in Crisis. Syst. Biol. 56(3). http://ezlab.zrc-sazu.si/uploads/big_files/Agnarsson&Kuntner2007SysBio.pdf
↑ ab Suarez, A. V., & Tsutsui, N. D. (2004). The value of museum collections for research and society. BioScience, 54(1), 66-74. http://bioscience.oxfordjournals.org/content/54/1/66.full
↑ abc Berendsohn, W.G., Güntsch, A., Hoffmann, N., Kohlbecker, A., Luther, K., Müller, A., 2011. Biodiversity information platforms: From standards to interoperability. Zookeys 150, 71–78. http://dx.doi.org/10.3897/zookeys.150.2166
↑ abcdefg Blagoderov, V., Kitching, I. J., Livermore, L., Simonsen, T. J., & Smith, V. S. (2012). No specimen left behind: industrial scale digitization of natural history collections. ZooKeys, (209), 133–146. doi:10.3897/zookeys.209.3178 http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3406472/
↑ abc Chavan, V. S., & Ingwersen, P. (2009). Towards a data publishing framework for primary biodiversity data: challenges and potentials for the biodiversity informatics community. BMC bioinformatics, 10(Suppl 14), S2. http://www.biomedcentral.com/1471-2105/10/S14/S2
↑ abcdef Heidorn, P. B. (2008). Shedding light on the dark data in the long tail of science. Library Trends, 57(2), 280-299. https://www.ideals.illinois.edu/bitstream/handle/2142/10672/heidorn.pdf?sequence=2
↑ ab Marshall, E. 2005. Will DNA Bar Codes Breathe Life Into Classification? Science. 307. http://barcoding.si.edu/BackgroundPublications%5CScience_Marshall_2005.pdf
↑ abcd McClain, C. (2011). The mass extinction of scientists who study species. Wired Science, 19(01). http://aberlentomo.fr/16_suppl_web_ento/mass_extinction_of_scientists_who_study_species.pdf
↑ abcd Lawrence, P. A. (2008). Lost in publication: how measurement harms science. Ethics in science and environmental politics, 8(1), 9-11.
↑ abc Costello, M. J. (2009). Motivating online publication of data. BioScience, 59(5), 418-427. http://bioscience.oxfordjournals.org/content/59/5/418.full
↑ Kratz, J., & Strasser, C. (2014). Data publication consensus and controversies. F1000Research, 3.
↑ abcdefghijklmnñ Payne, J. L., Smith, F. A., Kowalewski, M., Krause Jr, R. A., Boyer, A. G., McClain, C. R., Finnegan, S, Novack-Gottshall, PM & Sheble, L. (2012). A Lack of Attribution: Closing the Citation Gap Through a Reform of Citation and Indexing Practices. Taxon, 61(6), 1349-1351. http://www.ingentaconnect.com/content/iapt/tax/2012/00000061/00000006/art00030
↑ ab Boero, F. (2010). The study of species in the era of biodiversity: a tale of stupidity. Diversity, 2(1), 115-126. http://www.mdpi.com/1424-2818/2/1/115/pdf
↑ Zootaxa Impact Factor.http://www.mapress.com/zootaxa/support/impactfactor.htm
↑ FM Steiner, M Pautasso, H Zettel, K Moder, W Arthofer, BC Schlick-Steiner. 2015. A falsification of the citation impediment in the taxonomic literature. Syst Biol. syv026 http://sysbio.oxfordjournals.org/content/early/2015/05/04/sysbio.syv026.full.pdf
↑ abcdefghijklm Petr Pyšek, Philip E. Hulme, Laura A. Meyerson, Gideon F. Smith, James S. Boatwright, Neil R. Crouch, Estrela Figueiredo, Llewellyn C. Foxcroft, Vojtěch Jarošík, David M. Richardson, Jan Suda, and John R. U. Wilson (2013) Hitting the right target: taxonomic challenges for, and of, plant invasions AoB PLANTS (2013) 5: plt042 doi:10.1093/aobpla/plt042 first published online September 19, 2013 http://aobpla.oxfordjournals.org/content/5/plt042
↑ abc Minelli, A. (2003). The status of taxonomic literature. Trends in Ecology & Evolution, 18(2), 75-76.
↑ ab Krell, F. T. (2002). Why impact factors don't work for taxonomy. Nature, 415(6875), 957-957.
↑ Krell, F.T. (2010) Should editors influence journal impact factors? Learned Publishing, 23, 59–62.
↑ abcd Werner, Y. L. (2009). The aspiration to be good is bad: The “Impact Factor” hurts both science and society. Int J Sci Soc, 1, 99-105. http://ijy.cgpublisher.com/product/pub.187/prod.14
↑ abcdefghij Huang, X., Hawkins, B. A., Lei, F., Miller, G. L., Favret, C., Zhang, R., & Qiao, G. (2012). Willing or unwilling to share primary biodiversity data: results and implications of an international survey. Conservation Letters, 5(5), 399-406. http://onlinelibrary.wiley.com/doi/10.1111/j.1755-263X.2012.00259.x/full
↑ abcd Tautz, D.; Arctander, P., Minelli, A., Thomas, R. H., y Vogler, A. P. (2003). «A Plea for DNA Taxonomy» (pdf). Trends Ecol. Evol. 18 (2): 70-74. Archivado desde el original el 29 de septiembre de 2004. Consultado el 28 de agosto de 2007. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ ab Page, Roderic. 2009. Is taxonomy disadvantaged by standad measures of academic performance? (fue editor de Systematic Biology del 2004 al 2007) http://network.nature.com/groups/taxonomy/forum/topics/3983
Archivado el 31 de enero de 2016 en la Wayback Machine. Citado por Valdecasas (2011) Zootaxa.
↑ abcdefghijklmnñopqrstuvwx Carvalho, M. R., Ebach, M. C., Williams, D. M., Nihei, S. S., Trefaut Rodrigues, M., Grant, T., ... & Nelson, G. (2014). Does counting species count as taxonomy? On misrepresenting systematics, yet again. Cladistics, 30(3), 322-329.
↑ abc Valdecasas AG, Castroviejo S, Marcus LF (2000) Reliance on the citation index undermines the study of biodiversity. Nature 403(6771):698
↑ Valdecasas, A. G. (2011). An index to evaluate the quality of taxonomic publications. Zootaxa, 2925, 57-62. http://mapress.com/zootaxa/2011/f/zt02925p062.pdf
↑ abcd Thomas Borsch, P Hernández-Ledesma, WG Berendsohn, H Flores-Olvera, H Ochoterena, FO Zuloaga, S von Mering, N Kilian. 2015. An integrative and dynamic approach for monographing species-rich plant groups – Building the global synthesis of the angiosperm order Caryophyllales. Perspectives in Plant Ecology, Evolution and Systematics 17(4):284–300. Manuscrito aceptado: http://www.researchgate.net/profile/Patricia_Hernandez8/publication/277726151_An_integrative_and_dynamic_approach_for_monographing_species-rich_plant_groups__building_the_global_synthesis_of_the_angiosperm_order_Caryophyllales/links/55804f4408aea3d7096e4740.pdf
↑ ab Crisci, J. V. (2006). One-dimensional systematist: perils in a time of steady progress. Systematic botany, 31(1), 217-221. http://www.efn.unc.edu.ar/otros/bibliocentro/index_archivos/crisci.pdf
↑ abcdefg Wägele, H, A Klussmann-Kolb, M Kuhlmann,G Haszprunar, D Lindberg, A Koch, JW Wägele. 2011. The taxonomist - an endangered race. Fontiers Zool. http://www.biomedcentral.com/content/pdf/1742-9994-8-25.pdf
↑ abc Seifert KA, Crous PW, Frisvad JC (2008) Correcting the impact factors of taxonomic journals by appropriate citation of taxonomy (ACT). Persoonia 2008, 20:105. Según Wägele et al. 2011, propone unas prácticas "justas" para referenciar el trabajo del taxónomo, lo vuelve a citar entre otros que propusieron políticas para referenciar al taxónomo, para decir que fueron "largamente ignorados".
↑ van der Velde (2001) Taxonomists make a name for themselves. Nature.
↑ Krell, F-T. 2000 Impact factors aren't relevant to taxonomy. Nature.
Krell, F. T. (2002). Why impact factors don't work for taxonomy. Nature, 415(6875), 957-957.
Krell F-T. 2009. The poverty of citation databases: Data mining is crucial for fair metrical evaluation of research performance. BioScience 59: 6–7. http://bioscience.oxfordjournals.org/content/59/1/6.full.pdf+html
Krell, F.T. (2010) Should editors influence journal impact factors? Learned Publishing, 23, 59–62
↑ Ellis, J., 2002. Why impact factors don’t work for taxonomy. Nature 415, 957.
↑ Benítez, H. A. (2014). Is the Fever for High Impact a Disadvantage for Systematists?. Neotropical Entomology, 43(4), 295-297. http://www.researchgate.net/profile/Hugo_Benitez/publication/261366680_Is_the_Fever_for_High_Impact_a_Disadvantage_for_Systematists/links/00463536229cb45244000000.pdf
↑ abcdefghi McDade, L. A., Maddison, D. R., Guralnick, R., Piwowar, H. A., Jameson, M. L., Helgen, K. M., PS Herendeen, A Hill y Vis, M. L. (2011). Biology needs a modern assessment system for professional productivity. BioScience, 61(8), 619-625. http://bioscience.oxfordjournals.org/content/61/8/619.full
↑ Pyke, GH (2014) Evaluating the Quality of Taxonomic Publications: A Simple Alternative to Citations and Effort. BioScience first published online September 29, 2014 doi:10.1093/biosci/biu154 http://bioscience.oxfordjournals.org/content/early/2014/09/29/biosci.biu154.extract
↑ Páll-Gergely, B. (2014). Suggestions to improve the Taxonomy Index (T-Index) introduced by Valdecasas (2011). Zootaxa, 3780(2), 399-400. http://www.biotaxa.org/Zootaxa/article/download/zootaxa.3780.2.11/7374
↑ ab Venu, P., & Sanjappa, M. (2011). The impact factor and taxonomy. Current Science, 101(11), 1397-1397. http://repository.ias.ac.in/94703/1/The_impact_factor.pdf
↑ E Garfield (2001) Taxonomy is small, but it has its citation classics. Nature 413, 107 (13 September 2001) doi:10.1038/35093267 http://www.nature.com/nature/journal/v413/n6852/full/413107b0.html
↑ Como el taxónomo Liberty Hide Bailey alrededor de la década de 1930.
↑ ab Joppa, L.N., Roberts, D.L., Myers, N., Pimm, S.L., 2011. Biodiversity hotspots house most undiscovered plant species. Proc. Natl. Acad. Sci. U.S.A. 108, 13171–13176 http://dx.doi.org/10.1073/pnas.1109389108
↑ Von Staden, L., Raimondo, D., Dayaram, A., 2013. Taxonomic research priorities for the conservation of the South African flora. S. Afr. J. Sci. 109, 1–10. http://dx.doi.org/10.1590/sajs.2013/1182
↑ abcdefghijklm Costello, M. J., Michener, W. K., Gahegan, M., Zhang, Z. Q., & Bourne, P. E. (2013). Biodiversity data should be published, cited, and peer reviewed. Trends in Ecology & Evolution, 28(8), 454-461. http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.364.9572&rep=rep1&type=pdf
↑ abcde Carvalho, FA; D Filer, SS Renner. 2014. Taxonomy in the electronic age and an e-monograph of the papaya family (Caricaceae) as an example. Cladistics (2014)1-9. http://www.umsl.edu/~renners/Carvalho_E-monographing_Cladistics2014.pdf
↑ abc Patterson, D. J., Remsen, D., Marino, W. A., & Norton, C. (2006). Taxonomic indexing — extending the role of taxonomy. Systematic Biology, 55(3), 367-373.
↑ De Carvalho, M. R., Ebach, M. C., Williams, D. M., Nihei, S. S., Trefaut Rodrigues, M., Grant, T., … & Nelson, G. (2013). Does counting species count as taxonomy? On misrepresenting systematics, yet again. Cladistics.
↑ Cragin, M. H., Palmer, C. L., Carlson, J. R., & Witt, M. (2010). Data sharing, small science and institutional repositories. Philosophical Transactions of the Royal Society of London A: Mathematical, Physical and Engineering Sciences, 368(1926), 4023-4038. http://rsta.royalsocietypublishing.org/content/368/1926/4023.short
↑ abcd Costello, M. J., Vanhoorne, B., & Appeltans, W. (2015). Conservation of biodiversity through taxonomy, data publication, and collaborative infrastructures. Conservation Biology. http://www.researchgate.net/profile/Mark_Costello/publication/274723537_Conservation_of_biodiversity_through_taxonomy_data_publication_and_collaborative_infrastructures/links/552d0d7e0cf21acb0921248b.pdf
↑ ab Costello, M.J. and Vanden Berghe, E. (2006) ‘Ocean Biodiversity
Informatics’ enabling a new era in marine biology research and
management. Mar. Ecol. Prog. Ser. 316, 203–214 http://refhub.elsevier.com/S0169-5347(13)00109-2/sbref0005
↑ abcde Chavan, V., Penev, L., & Hobern, D. (2013). Cultural change in data publishing is essential. BioScience, 63(6), 419-420. http://bioscience.oxfordjournals.org/content/63/6/419.full
↑ Thomson Reuters. 2012. Thomson Reuters launches data citation index for discovering global data sets. Thomson Reuters. http://www.reuters.com/article/2012/10/17/idUS187519+17-Oct-2012+HUG20121017 (enlace roto disponible en Internet Archive; véase el historial y la última versión). citando a su vez a Thomson Reuters 2012. "Collaborative Science: Solving the Issues of Discovery, Attribution and Measurement in Data Sharing"
↑ Kratz, J. & Strasser, C. (2015b) Making data count. Sci. Data 2:150039 doi: 10.1038/sdata.2015.39 http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4550906/
↑ abc Force, M. M., & Robinson, N. J. (2014). Encouraging data citation and discovery with the Data Citation Index. Journal of computer-aided molecular design, 28(10), 1043-1048.
↑ Piwowar HA, Vision TJ. (2013) Data reuse and the open data citation advantage. PeerJ:e175 https://dx.doi.org/10.7717/peerj.175
A su vez cita a trabajos anteriores: "There is evidence that studies that make their data available do indeed receive more citations than similar studies that do not (Gleditsch, Metelits & Strand, 2003; Piwowar, Day & Fridsma, 2007; Ioannidis et al., 2009; Pienta, Alter & Lyle, 2010; Henneken & Accomazzi, 2011; Sears, 2011; Dorch, 2012)."
↑ Vines, T. H., Andrew, R. L., Bock, D. G., Franklin, M. T., Gilbert, K. J., Kane, N. C., Moore, J-S., Moyers, B. T., Renaut, S., Rennison, D. J., Veen, T. & Yeaman, S. (2013). Mandated data archiving greatly improves access to research data. The FASEB journal, 27(4), 1304-1308. http://www.fasebj.org/content/27/4/1304.full
↑ Canada’s Action Plan in Open Government. http://data.gc.ca/eng/ canadas-action-plan-open-government. Accessed 16 March 2014.
↑ The Open Government Partnership: Second Open Government National Action Plan for the United States of America. http://www.whitehouse.gov/sites/default/files/docs/us_national_action_plan_6p.pdf
Archivado el 13 de mayo de 2014 en la Wayback Machine.. Accessed 16 March 2014
↑ Research Councils UK. 28 de septiembre de 2015. Research > Open Access http://www.rcuk.ac.uk/research/openacess/ Research > Draft Concordat on Open Research Data http://www.rcuk.ac.uk/research/opendata/
↑ NSF Data Management Plan Requirements. https://www.nsf.gov/eng/general/dmp.jsp.
↑ Rausher, M. D., McPeek, M. A., Moore, A. J., Rieseberg, L. and Whitlock, M. C. (2010), DATA ARCHIVING. Evolution, 64: 603–604. doi: 10.1111/j.1558-5646.2009.00940.x
↑ abcd MC Whitlock, MA McPeek, MD Rausher, L Rieseberg y AJ Moore. 2010. Editorial: Data archiving. The American Naturalist 175(2):145-146 DOI: 10.1086/650340 Stable URL: http://www.jstor.org/stable/10.1086/650340
↑ Joint Data Archiving Policy (JDAP). http://datadryad.org/pages/jdap
↑ Bloom T, Ganley E, Winker M (2014) Data Access for the Open Access Literature: PLOS's Data Policy. PLoS Biol 12(2): e1001797. doi:10.1371/journal.pbio.1001797 http://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.1001797
↑ Parsons, M. A., & Fox, P. A. (2013). Is data publication the right metaphor?. Data Science Journal, 12(0), WDS32-WDS46. http://semanticommunity.info/@api/deki/files/29002/12_WDS-042.pdf
↑ Science 2007. Taxonomy, the early years. (Sobre Animal Base)
↑ The Biodiversity Heritage Library (BHL; http://biodiversitylibrary.org)
↑ JSTOR http://www.jstor.org
↑ Agosti, D, NF Johnson. 2002. Taxonomists need better access to published data. Nature.
↑ abc Causey, D., Janzen, D. H., Townsend, P. A., Vieglais, D., Krishtalka, L., Beach, J. H., & Wiley, E. O. (2004). Museum collections and taxonomy. Science, 305(5687), 1106b-1107b.
↑ De Carvalho, MR, FA Bockmann, DS Amorim, M de Vivo, M de Toledo-Piza, NA Menezes, JL de Figueiredo, RMC Castro, AC Gill, JD McEachran, LJV Compagno, RC Schelly, R Britz, JG Lundberg, RP Vari, G Nelson. 2004. Revisiting the Taxonomic Impediment. Science 307.
↑ ab Los proyectos http://www.zoosphere.net/ y https://www.idigbio.org/ que hacen fotos focus-stacking 3D de especímenes en museos. Erik Olsen, oct 19 2015. Museum specimens find new life online. The New York Times http://www.nytimes.com/2015/10/20/science/putting-museums-samples-of-life-on-the-internet.html
↑ JSTOR Global Plants.
↑ ab Blackmore, S. (2002). Biodiversity Update--Progress in Taxonomy. Science, 298(5592), 365-365. http://science.sciencemag.org/content/298/5592/365.short
↑ abcdefghi ZHI-QIANG ZHANG (2011) «Accelerating biodiversity descriptions and transforming taxonomic publishing: the first decade of Zootaxa.» Zootaxa 2896: 1-7 [2]
↑ ab «http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3261036/».
↑ Publicaciones de RDM Page.
↑ P ej IPNI
↑ abc Erwin, T; P Stoev, T Georgiev y L Penev. 2015. «ZooKeys 500: traditions and innovations hand-in-hand servicing our taxonomic community.» Zookeys. 2015; (500): 1–8. [3]
↑ abc Publicaciones de RDM Page
↑ Error en la cita: Etiqueta<ref>inválida;
no se ha definido el contenido de las referencias llamadasde Carvalho et al. 2007
↑ ab Raven, P. H. (2004). Taxonomy: where are we now?. Philosophical Transactions of the Royal Society B: Biological Sciences, 359(1444), 729. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1693354/pdf/15253358.pdf
↑ abc Hebert, P. D. N.; Cywinska, A., Ball, S. L. y De Waard, J. R. (2003). «Biological Identifications through DNA Barcodes» (pdf). Proc. R. Soc. Lond. Ser. B 270 (1512): 313-321. doi:10.1098/rspb.2002.2218. Consultado el 28 de agosto de 2007. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ De Carvallo et al. 2005. Revisiting the Taxonomic Impediment. Science.
↑ ab Ebach y Holdrege 2005. More taxonomy, not DNA barcoding. Bioscience.
↑ Will, Mishler, Wheeler. 2005. The Perils of DNA barcoding and the need for Integrative Taxonomy. Systematic Biology.
↑ abc Taylor, H. R., & Harris, W. E. (2012). An emergent science on the brink of irrelevance: a review of the past 8 years of DNA barcoding. Molecular Ecology Resources, 12(3), 377-388.
↑ abcde Taberlet, P., Coissac, E., Pompanon, F., Brochmann, C., & Willerslev, E. (2012). Towards next‐generation biodiversity assessment using DNA metabarcoding. Molecular ecology, 21(8), 2045-2050. https://www.researchgate.net/profile/Francois_Pompanon/publication/223968486_Towards_next-generation_biodiversity_assessment_using_DNA_metabarcoding/links/0912f500ff729af1be000000.pdf
↑ ab Hollingsworth PM, Forrest LL, Spouge JL et al. (2009) A DNA barcode for land plants. Proceedings of the National Academy of Sciences of the United States of America, 106, 12794–12797. http://www.pnas.org/content/106/31/12794.full?sid+38cee406-d425-4f58-8edc-bbcb2ac34bad
↑ INSDC (International Nucleotide Sequence Database Collaboration). http://www.insdc.org/ Un "shared pool" de datos moleculares como un acuerdo del DNA Data Bank of Japan (DDBI) con el European Molecular Biology Laboratory (EMBL) y el NCBI GenBank de Estados Unidos. Como comentado en Thessen y Patterson (2011).
↑ Barcoding of Life (BOLD) http://www.barcodeoflife.org/ (enlace roto disponible en Internet Archive; véase el historial y la última versión).
Barcode of Life Data System http://www.boldsystems.org/views/login.php
↑ Savolainen, V.; R. S. Cowan, A. P. Vogler, George K. Roderick, y Richard Lane (2005). «Towards writing the encyclopaedia of life: an introduction to DNA barcoding» (pdf). Phil. Trans. R. Soc. B: 1-7. doi:10.1098/rstb.2005.1730. Archivado desde el original el 12 de octubre de 2008. Consultado el 9 de mayo de 2009. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ Consortium for the Barcode of Life. (CBOL) http://www.barcoding.si.edu/
↑ Siddall, M. E.; P. Trontelj, S. Y. Utevsky, M. Nkamany y K. S. Macdonald (2007). «Diverse molecular data demonstrate that commercially available medicinal leeches are not Hirudo medicinalis» (pdf). Proc. R. Soc. B 274 (1617): 1481-1487. Consultado el 9 de mayo de 2009. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ DeSalle, R., Egan, M. G., & Siddall, M. (2005). The unholy trinity: taxonomy, species delimitation and DNA barcoding. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 360(1462), 1905-1916. http://rstb.royalsocietypublishing.org/content/360/1462/1905.short
↑ Godfray HCJ, Mayo SJ, Scoble MJ. 2008b. Pragmatism and rigour can coexist in taxonomy. Evolutionary Biology 35:309 –311. http://link.springer.com/article/10.1007/s11692-008-9041-8
↑ PH Raven, 2002. Science, sustainability and the human prospect. Science 297:954-958.
↑ Jablonski, N. G. (2005). Exploring Biodiversity: Past, Present, and Future. PROCEEDINGS-CALIFORNIA ACADEMY OF SCIENCES, 56, 9.
↑ Joppa, LN; P Visconti, CN Jenkins, SL Pimm. 2013. Achieving the Convention on Biological Diversity's Goals for Plant Conservation. Science 341(6150):1100-1103. DOI: 10.1126/science.1241706 http://www.sciencemag.org/content/341/6150/1100.abstract
↑ Myers, N., Mittermeier, R.A. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853–859.
↑ Derek P. Tittensor, Matt Walpole, Samantha L. L. Hill, Daniel G. Boyce, Gregory L. Britten, Neil D. Burgess, Stuart H. M. Butchart, Paul W. Leadley, Eugenie C. Regan, Rob Alkemade, Roswitha Baumung, Céline Bellard, Lex Bouwman, Nadine J. Bowles-Newark, Anna M. Chenery, William W. L. Cheung, Villy Christensen, H. David Cooper, Annabel R. Crowther, Matthew J. R. Dixon, Alessandro Galli, Valérie Gaveau, Richard D. Gregory, Nicolas L. Gutierrez, Tim L. Hirsch, Robert Höft, Stephanie R. Januchowski-Hartley, Marion Karmann, Cornelia B. Krug, Fiona J. Leverington, Jonathan Loh, Rik Kutsch Lojenga, Kelly Malsch, Alexandra Marques, David H. W. Morgan, Peter J. Mumby, Tim Newbold, Kieran Noonan-Mooney, Shyama N. Pagad, Bradley C. Parks, Henrique M. Pereira, Tim Robertson, Carlo Rondinini, Luca Santini, Jörn P. W. Scharlemann, Stefan Schindler, U. Rashid Sumaila, Louise S.L. Teh, Jennifer van Kolck, Piero Visconti, y Yimin Ye. 2014. A mid-term analysis of progress toward international biodiversity targets. Science 10 October 2014: 346 (6206), 241-244. Published online 2 October 2014 [DOI:10.1126/science.1257484] http://www.sciencemag.org/citmgr?gca=sci%3B346%2F6206%2F241
Archivado el 24 de septiembre de 2015 en la Wayback Machine.
↑ Peterson, AT; A. Navarro- Sigüenza (1999). «Alternate species concepts as bases for determining priority conservartion areas». Conservation Biology 13: 427-431. DOI 10.1046/j.1523-1739.1999.013002427.x. Consultado el 17 de julio de 2009. La referencia utiliza el parámetro obsoleto|coautores=(ayuda)
↑ Harcourt, A.H. et al. (2002) Rarity, specialization and extinction in primates. J. Biogeogr. 29, 445–456
↑ Guralnick, R., Conlin, T., Deck, J., Stucky, B. J., & Cellinese, N. (2014). The trouble with triplets in biodiversity informatics: a data-driven case against current identifier practices. PloS one, 9(12), e114069. http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0114069
↑ GBIF http://www.gbif.org
↑ «http://www.inaturalist.org/».
↑ «http://ebird.org/content/ebird/».
↑ «http://www.gbif.org/whatisgbif».
↑ Secies 2000 http://www.sp2000.org
↑ Encyclopedia of Life http://eol.org
↑ «http://www.itis.gov».
↑ Wilson, E. O. (2003). The encyclopedia of life. Trends in Ecology and Evolution, 18(2), 77-80.
↑ Stuart, S. N., Wilson, E. O., McNeely, J. A., Mittermeier, R. A., & Rodríguez, J. P. (2010). The barometer of life. Science, 328(5975), 177-177.)
↑ Tree of Life http://www.tolweb.org/tree/phylogeny.html
↑ «http://www.tolweb.org/tree/home.pages/history.html».
↑ «http://www.tolweb.org/tree/home.pages/toleol.html».
↑ Tree of Life Growth Monitor.
http://www.tolweb.org/tree/home.pages/growth.html
↑ Costello, M. J., Appeltans, W., Bailly, N., Berendsohn, W. G., de Jong, Y., Edwards, M., ... & Bisby, F. A. (2014). Strategies for the sustainability of online open-access biodiversity databases. Biological Conservation, 173, 155-165. http://www.sciencedirect.com/science/article/pii/S0006320713002747
↑ abcd Greg Miller (2005) Linnaeus's Legacy Carries On. Science 307(5712). http://www.sciencemag.org/content/307/5712/1038.1.summary
↑ Berger, J. K. (2005). Mission possible: ALL Species Foundation and the call for discovery. PROCEEDINGS-CALIFORNIA ACADEMY OF SCIENCES, 56, 114.
↑ Wilson, E.O. 2003. The encyclopedia of life. Trends in Ecology and Evolution 18:77–80
↑ May, R. M., & Harvey, P. H. (2009). Species uncertainties. Science, 323(5915), 687-687. http://www.sciencemag.org/content/323/5915/687.full
↑ Ariño AH. (2010) Approaches to estimating the universe of natural History collections data. Biodiversity Informatics 7: 81-92
↑ Zootaxa (2015) Impact Factor http://www.mapress.com/zootaxa/support/impactfactor.htm
↑ ab ZHI-QIANG ZHANG, HANS-JOACHIM ESSER & MAARTEN J.M. CHRISTENHUSZ (2013) Editorial. «Phytotaxa 100: The rise of a major journal in systematic and taxonomic botany.» Phytotaxa 100 (1): 1–5 http://www.mapress.com/phytotaxa/content/2013/f/pt00100p005.pdf
↑ ZHI-QIANG ZHANG, MAARTEN J.M. CHRISTENHUSZ, HANS-JOACHIM ESSER, MARK W. CHASE, MARIA S. VORONTSOVA, HEATHER LINDON, ALEXANDRE MONRO & H. Thorsten Lumbsch. 2014. «The making of world’s largest journal in systematic botany.» Phytotaxa 191 (1): 001–009 http://www.mapress.com/phytotaxa/content/2014/f/pt00191p009.pdf
↑ ab Miller, J. A., Georgiev, T., Stoev, P., Sautter, G., & Penev, L. (2015). Corrected data re-harvested: curating literature in the era of networked biodiversity informatics. Biodiversity data journal, (3). http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4304254/
↑ Alessandro Minelli (2013) «Zoological nomenclature in the digital era.» Front Zool. 2013; 10: 4. [4]
↑ Minelli, A. (2001) Short notes and reviews Zoological nomenclature – reflections on the recent past and ideas for our future agenda. Bulletin of Zoological Nomenclature, 58, 164–169.
↑ Christina Flann. 2011. BioCode: Third time lucky? Zootaxa 2874
↑ Orchard, A.F., W.R. Anderson, M.G. Gilbert, D. Sebsebe, W.T. Stearn, and E.G. Voss. 1996. Harmonized bionomenclature – a recipe for disharmony. Taxon 45: 287-290.
↑ R. K. Brummitt. 1997. The BioCode is unnecessary and unwanted. Systematic Botany 22: 182-186. También ver Brummitt, R.K. 1996. "Quite happy with the present code, thank you." En: J.L. Reveal, ed. Proceedings of a mini-symposium on biological nomenclature in the 21st century. University of Maryland: http://www.plantsystematics.org/bees/96sym.html (enlace roto disponible en Internet Archive; véase el historial y la última versión)..
↑ Delprete, P. 1996. "COMMENTARY 1: Biological Nomenclature". En: J.L. Reveal, ed. Proceedings of a mini-symposium on biological nomenclature in the 21st century. University of Maryland: http://www.plantsystematics.org/bees/96sym.html (enlace roto disponible en Internet Archive; véase el historial y la última versión)..
↑ Werner Greuter (2011) BioCode 2011. Explanatory prologue. http://www.bionomenclature.net/biocode2011.html
↑ McNeill, John y NJ Turland. 2011. Major changes to the Code of Nomenclature - Melbourne, July 2011. Taxon 60(5)October 2011:1495-1497. http://www.seaweed.ie/nomenclature/pdf/taxon_60_major_changes_xviii_ibcm.pdf
↑ «Código Internacional para la Nomenclatura de Bacterias» (en inglés). Consultado el 28 de agosto de 2007.
↑ International Commission on Zoological Nomenclature (2012) Amendment of Articles 8, 9, 10, 21 and 78 of the International Code of Zoological Nomenclature to expand and refine methods of publication. Zootaxa, 3450, 1–7.
↑ Zoobank http://www.zoobank.org/About
↑ Vincent Robert et al. (2013) MycoBank gearing up for new horizons. IMA Fungus. 2013 Dec; 4(2): 371–379.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3905949/
↑ de Queiroz, K. 1997. The Linnaean hierarchy and the evolutionization of taxonomy, with emphasis on the problem of nomenclature. Aliso 15: 125-144.
↑ International Commision of Zoological Nomenclature. Visitado en agosto 2013. FAQ. "Should I use PhyloCode?" [5]
↑ abcdefg Nixon, K.C., Carpenter, J.M., Stevenson, D.W., 2003. The PhyloCode is fatally flawed, and the “Linnaean system” can easily be fixed. Bot. Rev. 69, 111-120. http://www.botany.wisc.edu/courses/botany_940/05PhyloCode/papers/NixonCarpen2003.pdf Paper que retoma a
Nixon, K. C., y J.M. Carpenter. 2000. On the Other "Phylogenetic Systematics". Cladistics 16, 298–318 (2000) doi:10.1006/clad.2000.0137 http://155.97.32.9/~mhaber/Documents/Course%20Readings/NixonCarpenter_Clad2000.pdf
Archivado el 7 de septiembre de 2015 en la Wayback Machine.
↑ abc Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
↑ ab Potter, D., Eriksson, T., Evans, R. C., Oh, S., Smedmark, J. E. E., Morgan, D. R., ... & Campbell, C. S. (2007). Phylogeny and classification of Rosaceae. http://biology.umaine.edu/Amelanchier/Rosaceae_2007.pdf Plant Systematics and Evolution, 266(1-2), 5-43.
Error en la cita: La etiqueta <ref> con nombre "Collins y Cruickshank 2013" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Sokal 1975" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Pires y Marinoni 2010" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Vermeij 2005" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Wilson 1985" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Hey et al. 2009" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Valdecasas et al. 2008" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Güntsch et al. 2013" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Godfray et al. 2007" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Wheeler y Meier 2013" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Donoghue y Kandereit 1992" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Buffon 1749" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Dobzhansky 1937" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Simpson 1951" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Adl et al. 2012" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Ruggiero et al. 2015" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Donoghue y Cantino 1988" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Cronquist 1987" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Stuessy 1983" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Reveal 2012" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Gunn 2009" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Huffman y Wallace 2012" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Ung et al. 2010" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Darwin 1859" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Stevens 1994" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta <ref> con nombre "Minelli 2008" definida en <references> no se utiliza en el texto precedente.
Error en la cita: La etiqueta
<ref> con nombre "McNeill 2014" definida en <references> no se utiliza en el texto precedente.
Bibliografía
Judd, W. S.; Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. (2007). Plant systematics: a phylogenetic approach, Third Edition (Sinauer Axxoc edición). USA. ISBN 0-87893-403-0. La referencia utiliza el parámetro obsoleto|coautores=(ayuda) Capítulos de taxonomía general y teórica: 1, 2, 3, 5, 6, Apéndice I, Apéndice II.
- Cavalier-Smith, T. (2010). Deep phylogeny, ancestral groups and the four ages of life. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1537), 111-132.
Enlaces externos
 Wikcionario tiene definiciones y otra información sobre Taxonomía.
Wikcionario tiene definiciones y otra información sobre Taxonomía.
- El Diccionario de la Real Academia Española tiene una definición para Taxonomía.


