Aile (anatomie des insectes)
Cet article ne cite pas suffisamment ses sources (septembre 2009).
Si vous disposez d'ouvrages ou d'articles de référence ou si vous connaissez des sites web de qualité traitant du thème abordé ici, merci de compléter l'article en donnant les références utiles à sa vérifiabilité et en les liant à la section « Notes et références »
En pratique : Quelles sources sont attendues ? Comment ajouter mes sources ?
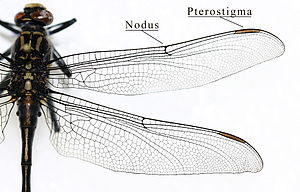
Ailes d'une libellule (insecte odonate).

Détail d'une aile de papillon (insecte lépidoptère).
L'aile de l'insecte est une expansion tégumentaire de l'exosquelette de l'insecte qui lui permet de voler. Elle n'est absolument pas homologue à l'aile des oiseaux.
Les ailes sont présentes dans la sous-classe des ptérygotes (cf. grec πτέρυξ/-γ, « ptéryx » / « aile ») et absentes chez les aptérygotes. Elles se mettent en place lors de la dernière mue. Lorsque l'insecte sort de son exuvie, ses ailes sont toutes froissées et repliées sur elles-mêmes, c'est la pression de l'hémolymphe qui contribue à les rigidifier et à leur donner leur forme définitive.
La forme des ailes constitue une clé importante pour la détermination des ordres. Ceux-ci ont souvent comme nom un mot avec un suffixe « -ptère ».
Sommaire
1 Morphologie : généralités et exceptions
2 Structure
2.1 Veines ou nervures
2.2 Cellules
2.2.1 Système de nommage
2.3 Poils et écailles
2.4 Superhydrophobie
2.5 Autres structures
3 Origine et évolution
4 Vol
5 Adaptations
5.1 Pour l'équilibrage
5.2 Pour la protection
5.3 Autres adaptations
6 Génétique
7 Notes et références
8 Bibliographie
9 Voir aussi
9.1 Articles connexes
9.2 Liens externes
Morphologie : généralités et exceptions |
Les ailes sont insérées sur le deuxième et le troisième segment thoracique (le mésothorax et le métathorax), pas sur le premier segment (prothorax) chez qui le gène Hox réprime leur formation. Ce ne sont pas des « appendices » d'un point de vue technique, puisque les insectes n'ont qu'une seule paire d'appendices par segment : les pattes. Les ailes sont renforcées par un certain nombre de veines longitudinales, qui possèdent souvent des interconnexions formant des « cellules » fermées dans la membrane (les exemples extrêmes sont à trouver parmi les odonates et les névroptères). Le dessin résultant des fusions et interconnexions des veines des ailes est souvent un caractère de détermination des différentes lignées évolutives et peut être utilisé pour l'identification de la famille ou même du genre dans de nombreux ordres d'insectes.
Les ailes totalement fonctionnelles ne sont présentes que chez les adultes. La seule exception est l'ordre des éphéméroptères chez qui l'avant-dernière étape larvaire subimago possède des ailes bien développées et fonctionnelles, dont l'animal se sépare lors de la mue finale. Chez les insectes qui ont un développement post-embryonnaire avec larves et nymphes (Diptères, Lépidoptères, Hyménoptères…) les ailes ne sont pas visibles chez la larve mais sont présents à l'état de « disque imaginal ».
Les insectes ont généralement deux paires d'ailes, sauf chez les Diptères où l'aile du 3e segment est remplacée par des balanciers, chez les strepsiptères où c'est l'aile du 2e segment qui est remplacée, chez les coléoptères où l'aile du 2e segment est transformée en élytre. Les membracidae ont exceptionnellement 3 paires d'ailes, la 3e étant modifiée en « casque ».
Les ailes peuvent être totalement absentes dans certains taxons des ptérygotes, comme chez les puces ou les poux. Bien que leurs ancêtres aient été ailés, le mode de vie parasitaire (dans la fourrure des animaux) de ces insectes fait que la présence d'ailes constituerait une gêne. Au cours de l'évolution, ils ont donc perdu ces appendices, mais la constitution de leur thorax indique qu'ils dérivent bien d'ancêtres ailés.
Les ailes peuvent n'être présentes que chez un seul sexe (souvent le mâle) dans certains groupes comme les Mutillidae et les Strepsiptera, ou perdues sélectivement chez les « travailleurs » des insectes sociaux tels que les fourmi et les termites. Rarement, la femelle est ailée et le mâle non, comme chez les guêpes des figues. Dans certains cas, les ailes sont produites seulement à des moments particuliers du cycle de vie, comme durant la phase de dispersion des Aphidoidea. Au-delà de la simple présence/absence des ailes, la structure et la coloration varie souvent au sein d'une même espèce par polymorphisme génétique.
Au repos, les ailes peuvent être tenues à plat, ou pliées un nombre de fois défini selon un motif spécifique ; en général, ce sont les ailes postérieures qui sont pliées, mais dans quelques groupes comme les guêpes Vespidae, ce sont les ailes antérieures.
Structure |

La nervation d'une aile de papillon (insecte lépidoptère).

Coupe transversale d'une nervure d'une aile.
L’aile est une membrane cuticulaire formée par la juxtaposition de deux couches épidermiques. Cette structure délicate représente 0.5 à 5 % de la masse totale corporelle de l'insecte[1].
Veines ou nervures |
Un schéma de nervation primitive hypothétique est souvent utilisé pour décrire les dessins de nervation des espèces existantes. Cet archétype primitif est appelé « archedictyon ».
Les nervures (ou veines) sont des structures creuses tubulaires formées par le couplage des parois supérieures et inférieures de l'aile. Les plus grosses veines sont des voies de passage de l'hémolymphe, des systèmes trachéaux et nerveux (neurones des organes sensoriels)[1]. L'aile de l’insecte, contrairement à une croyance répandue, n'est donc pas un organe mort et sec[2] : les trachées apportent de l'oxygène, l'hémolymphe fournit des nutriments et des hormones nécessaires au métabolisme des neurones, et de l'eau pour maintenir l'élasticité et la dureté de l'aile. Les organes sensoriels sont principalement des mécanorécepteurs qui interviennent dans le contrôle du mouvement des ailes et de la stabilisation du corps en vol (détection des distorsions de la cuticule). Chez certains insectes, des mécanorécepteurs sensibles aux vibrations pour détecter des prédateurs et servant à la communication intraspécifique. Il existe également des chimiorécepteurs (en lien avec le toilettage[3], le contrôle des qualités nutritives des sources du nectar[4] et les thermorécepteurs (rotection contre la surchauffe)[5].
Les nervures plus petites sont principalement des structures rigidifiantes formées de chitine. Le schéma de la nervation est extrêmement variable et peut être utile pour l'identification des espèces, par exemple pour distinguer les libellules des genres Cordulia et Somatochlora.
La circulation de l'hémolymphe dans la lumière des nervures des ailes, étudiée dès 1831 par le savant Carl Gustav Carus[6], est bien définie : l'hémolymphe entre par les nervures antérieures et sort par les nervures postérieures[7]. L'hémolymphe est mue par des organes pulsatiles thoraciques (« cœurs » accessoires pairs ou impairs)[2]. La physiologie du cœur chez certains Lépidoptères, Coléoptères et Diptères[8] présente une particularité : la circulation n'est pas circulaire mais alternée dans chaque nervure, grâce au renversement des battements cardiaques[9].
Les veines procurent à la fois de la rigidité et de la flexibilité à l'aile, ce qui permet à l'insecte de voler. De façon plus importante, la membrane de l'aile entre les nervures est très souvent ondulée, distordue ou plissée afin de donner encore plus de force à l'aile, ainsi que de changer la forme de la section de l'aile, d'une forme de feuillet plat vers une forme d'aile d'avion.
Cellules |
Les cellules sont les espaces de membrane formés entre les connexions des veines. Elles peuvent être « ouvertes » et s'étendre jusqu'au bord de l'aile ou « fermées » et être délimitées seulement par des veines.
Système de nommage |
Les veines et les cellules sont nommées d'après le système Comstock-Needham (1898).
Poils et écailles |
Les ailes des insectes peuvent être recouvertes de poils ou d'écailles.
Il s'agit du caractère qui définit l'ordre des lépidoptères (les papillons) chez qui les ailes sont recouvertes d'écailles colorées, ce sont des soies aplaties. Le terme lépidoptère vient du mot grec « lepidos » qui signifie « écaille ». Ce sont ces écailles qui donnent leurs couleurs aux ailes des papillons. Certains d'entre eux (genre Morpho, par exemple) ont des couleurs irisées qui viennent de la diffraction de la lumière sur des sillons des écailles, bien que leurs ailes ne contiennent aucun pigment.

Poils sur l'aile d'une mouche.

Écailles vues en plus fort grossissement.

Couleur bleue irisée des ailes d'un Morpho.



Superhydrophobie |
Les ailes d'insecte sont revêtues de plots micro- et nanostructurés, formant une surface superhydrophobe (lorsque des gouttes d'eau tombent dessus, elles roulent et évacuent la poussière sans mouiller ces surfaces), auto-nettoyante et aux propriétés anti-microbiennes[10].
Autres structures |
On appelle « Ptérostigma » un épaississement du bord antérieur des ailes, et « tégula », les petites écailles qui protègent la base des ailes antérieures.
Origine et évolution |
Au cours du Carbonifère inférieur, les insectes Ptérygotes ont commencé à voler il y a environ 350 millions d'années[11]. La rareté des fossiles à cette période ne permet pas de dire précisément comment et pourquoi les ailes des insectes se sont développées mais ces fossiles présentent sur tous les segments thoraciques et abdominaux des excroissances homologues d'ailes en série qui ont été réprimées par l'expression des gène Hox au cours de l'évolution, à l'exception des mésothorax et métathorax[12].
Les deux principales hypothèses pour expliquer l'origine des ailes sont :
- Expansion des tergites du thorax en palettes qui auraient aidé les premiers hexapodes à se déplacer dans les arbres. On retrouve en effet des coussinets aplatis sur des espèces fossiles. Ces coussinets auraient d’abord servi à réguler la température ou servir à glisser avant de devenir de véritables ailes[13].
- Transformation de branchies à partir de larves aquatiques : un fossile de nymphe d'un éphémère découvert en 1981 en Tchécoslovaquie possédait des branchies planes sur chaque segment abdominal et aussi trois paires d'ailes atrophiées. La biologiste évolutionniste Jarmila Kukalova-Peck en déduit que les ailes se sont développées à partir de branchies présentes sur les pattes des ancêtres des plécoptères[14].
La phylogénie moléculaire (selon le modèle de l'horloge moléculaire) suggère que l'apparition des ailes au cours de l'évolution ait pu avoir lieu au moment de la croissance spectaculaire des plantes dans les forêts du Dévonien, pour faciliter la redescente sur le sol de ces insectes se nourrissant à l'extrémité des branches[15].
Vol |
Le vol de l'insecte peut être extrêmement rapide, manœuvrant et versatile. Ces performances de vol sont possibles grâce aux changements de formes, un contrôle très fin et variable des mouvements de l'aile. Différents mécanismes de vol se rencontrent à travers les ordres.
Adaptations |
Plusieurs ordres d'insectes ont des ailes spécialement adaptées.
Pour l'équilibrage |
Chez les diptères (mouches, moustiques…), la paire d'aile postérieure est réduite à des haltères, qui aide l'insecte à ressentir son orientation et mouvement, et à améliorer son équilibre en vol en agissant à la manière de gyroscopes.
Chez les Strepsiptères, c'est la paire d'aile antérieure des mâles qui sont réduites pour former des haltères. Les femelles sont aptères.
Pour la protection |
Chez les coléoptères (scarabées), la paire d'aile antérieure est endurcie pour former des élytres qui protègent les ailes postérieures délicates, celles-ci sont repliées en dessous au repos. Les élytres ne servent pas au vol, seules les ailes postérieures des coléoptères lui servent à voler. Lorsque le coléoptère s'apprête à s'envoler, les élytres se soulèvent et les ailes postérieures sont tirées vers l'avant par les muscles thoraciques et sont dépliées. Le mécanisme qui sous-tend le déploiement de l'aile fait l'objet de débats : libération de l'énergie élastique stockée dans les structures contenant de la résiline (en)[16], rôle crucial de la pression de l'hémolymphe pour les déplier[17].
Chez certaines espèces, telles les carabes, les élytres sont soudés l'une à l'autre (les ailes postérieures ayant disparu), ce qui a fait perdre la faculté de voler à ces insectes. Cette incapacité a été remplacée par le développement de longues pattes puissantes qui leur assurent une marche rapide[18].

Élytres ouverts d'un hanneton.

Élytres soudés d'un carabe.


Chez les dermaptères, orthoptères (sauterelles, criquets), dictyoptères (blattes, mantes), on appelle « tegmen » (pluriel « tegmina ») les ailes antérieures renforcées, qui ne sont pas utilisées pour voler. On appelle aussi parfois ces ailes « élytres », ou « pseudoélytres ».
Chez les hémiptères, les ailes antérieures peuvent être renforcées, mais dans une moindre mesure que chez les coléoptères. Par exemple, la partie antérieure de l'aile de devant de la punaise est endurcie, tandis que la partie postérieure est membraneuse. On appelle ces ailes « hemelytron » (pluriel « hemelytra »). Elles sont trouvées dans le sous ordre des hétéroptères ; les ailes des homoptères, telles celles de la cigale, sont entièrement membraneuses.

Hémelytres d'une punaise verte.

Chez un certain nombre d'autres ordres, les ailes antérieures peuvent occasionnellement être modifiées pour la protection, et ceci se passe habituellement en conjonction avec la perte ou la réduction des ailes postérieures (par exemple chez les insectes ne volant pas). De façon similaire les individus non-volants des ordres précédents n'ont absolument pas d'ailes postérieures.
Autres adaptations |
- Les hyménoptères, dont le nom provient du Grec hymên, « mariage », et ptéron, « aile », du fait qu'il existe des crochets sur la nervure antérieure de l'aile postérieure permettant à celle-ci d'être couplée avec les ailes antérieures.
- Dans l’ordre des orthoptères les ailes sont modifiées pour aider à la production sonore. Chez les ensifères ceci est accompli par le frottement du haut des ailes, qui a un instant de crissement comme structure, contre l’une à l’autre pendant que les fémurs postérieurs sont frottés contre les ailes chez les caelifères (voir aussi stridulation). Chez peu de sauterelles, la structure de la production sonore fonctionne uniquement quand les ailes sont battues, avec les ailes antérieurs et les ailes postérieures qui frappent l’une et l’autre, et chez certains lépidoptères (par exemple, les hamadryas), le son est produit par les ailes antérieures frappant l’une et l’autre au sommet du trait.
- Certaines espèces utilisent leurs ailes pour la thermorégulation comportementale (voir par exemple la posture dorsale et latérale des papillons). La contraction des muscles alaires du vol réchauffe l'hémolymphe du thorax qui est diffusée vers l’abdomen où elle se refroidit[19].
- Certaines espèces de coléoptères de la famille des ténébrions dans le désert du Namib ont des élytres qui agissent comme des surfaces comme des voiles pour condenser et avoir une arête pour dévier l’eau vers leur bouche.
- Des coléoptères aquatiques tels le dytique (genre Dytiscus) utilisent l'espace entre leurs élytres et leur abdomen pour emprisonner de l'air lors de leur déplacement sous l'eau.
- Chez certains insectes sociaux, tels les termites ou les fourmis, seuls les individus des castes royales sont munies d'ailes. Ils s'en servent lors de leur parade nuptiale, puis s'en débarrassent au moment de fonder une nouvelle colonie.
Génétique |
Chez la mouche drosophile (Drosophila melanogaster, Diptère) :
- Il existe une mutation donnant des ailes vestigiales, c’est-à-dire des ailes « embryonnaires », non fonctionnelles.
- La paire d'aile différenciée en haltères des diptères se fait sous le contrôle génétique d'un gène appelé « Ultrabithorax ». La mutation de ce gène aboutit à un adulte avec 4 ailes au lieu de 2.
- le gène « Arc » donne des ailes en arc.
Notes et références |
(en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « insect wing » (voir la liste des auteurs).
(en) Günther Pass, « Beyond aerodynamics: The critical roles of the circulatory and tracheal systems in maintaining insect wing functionality », Arthropod Structure & Development, vol. 47, no 4, 2018, p. 391-407.
Alain Fraval, « Le système circulatoire », Insectes, no 166, 2012, p. 29 (lire en ligne).
(en) A. Yanagawa, A.M. Guigue, F. Marion-Poll, « Hygienic grooming is induced by contact chemicals in Drosophila melanogaster », Front. Behav. Neurosci., 8 (2014), p. 254
(en) H. Raad, J.-F. Ferveur, N. Ledger, M. Capovilla, A. Robichon, « Functional gustatory role of chemoreceptors in Drosophila wings », Cell Rep., 15 (2016), p. 1442-1454
H. Schmitz, L.T. Wasserthal, « Antennal thermoreceptors and wing thermosensitivity of heliotherm butterflies: their possible role in thermoregulatory behavior », J. Insect Physiol., 39 (1993), p. 1007-1019
(de) C.G. Carus, « Fernere Untersuchungen über Blutlauf in Kerfen », Nova Acta Physico. Med. Acad. Caes. Leop. Carol., 15 (1831), p. 1-18
(en) J.W. Arnold, « Blood circulation in the insect wings », Mem. Entomol. Soc. Can., 38 (1964), p. 5-60
(en)L.T. Wasserthal, « Interaction of circulation and tracheal ventilation in holometabolous insects », Adv. Insect Physiol., 26 (1996), p. 297-351
(en) J.H. Gerould, « History of the discovery of periodic reversal of heartbeat in insects », Biol. Bull, 56 (1929), p. 215-225
(en) Doyoung Byun, Jongin Hong, Saputra, Jin Hwan Koa, Young Jong Leea, Hoon Cheol Park, Bong-Kyu Byund, Jennifer R. Lukesb, « Wetting Characteristics of Insect Wing Surfaces », Journal of Bionic Engineering, vol. 6, no 1, 2009, p. 63-70 (DOI 10.1016/S1672-6529(08)60092-X).
(en) David Grimaldi, Michael S. Engel, Evolution of the Insects, Cambridge University Press, 2005, p. 153-154.
(en) T. Ohde, T. Yaginuma et T. Niimi, « Insect Morphological Diversification Through the Modification of Wing Serial Homologs », Science, 28 février 2013(DOI 10.1126/science.1234219)
(en) David Grimaldi, Michael S. Engel, Evolution of the Insects, Cambridge University Press, 2005, 772 p. (ISBN 0521821495)
Jarmilak Ukalova-Peck, Origin of the insect wing and wing articulation from the arthropodan leg, Department of Geology, Carleton University, Ottawa, Ont., Canada KlS 5B6, 17 mai 1982
À la recherche de l'origine des insectes Émission Continent sciences sur France Culture le 6 septembre 2010
(en) F. Haas, S. Gorb, R. Blickhahn, « The function of resilin in beetle wings », Proc. R. Soc. Lond. B, 267 (2000), p. 1375-1381
(en), J. Sun, W. Wu, M. Ling, J. Tong, L. Ren, « Fluid analysis of vein of beetle hindwing during unfolding action », Int. J. Heat Mass Transfer, 101 (2016), p. 379-386
Dominique Mariau, L'univers fascinant des insectes, L'Harmattan, 2012, p. 25.
Alain Fraval, « Le système circulatoire », Insectes, no 166, 2012, p. 28 (lire en ligne).
Bibliographie |
- (en) Charles A. Triplehorn, Norman F. Johnson et Donald Joyce Borror, Borror and DeLong's introduction to the study of insects, Thomson Brooks/Cole, 2005, 7e éd., 864 p. (ISBN 0-03-096835-6, OCLC 55793895)
Maurice Roth, Initiation à la morphologie, la systématique et la biologie des insectes, vol. 23, Paris, ORSTOM, coll. « Initiations-documentations techniques », 1980, 213 p. (ISBN 2-7099-0327-X et 978-2709903271, présentation en ligne, lire en ligne) [PDF]
Voir aussi |
Articles connexes |
- Vol de l'insecte
- Haltère de l'insecte
- Ocelle
Formule alaire, technique de biométrie
- Brachyptère et aptères dans le Wiktionnaire
Liens externes |
(en) « Cours sur l'aile de l'insecte », sur Université de Caroline du Nord
- Alain Ramel, « Petit cours illustré d'entomologie - Les ailes », sur Le Monde des Insectes
 Portail de l’entomologie
Portail de l’entomologie  Portail de l’anatomie
Portail de l’anatomie


